
Antropogeneza (gr. ánthrōpos – człowiek, génesis – pochodzenie) – filogeneza człowieka, czyli pochodzenie, droga rozwojowa i procesy biologiczne, które doprowadziły do powstania anatomicznie nowoczesnego człowieka rozumnego (Homo sapiens), wyjaśniane na gruncie teorii ewolucji w oparciu o osiągnięcia antropologii, filogenetyki molekularnej i antropolingwistyki.
Jedną z proponowanych hipotez jest ta autorstwa polskich naukowców, Konrada Fiałkowskiego i Tadeusza Bielickiego, dotycząca powstania mózgu ludzkiego dzięki wcześniejszej adaptacji mózgu hominidów do warunków stepowiejącego środowiska tropikalnej sawanny Afryki Wschodniej (późny pliocen – wczesny i środkowy plejstocen). Warunki klimatyczne i ostra konkurencja z silniejszymi drapieżnikami wymusiły stosowanie specyficznej techniki polowania – metody długotrwałego biegu, nieprzerywanego mimo sygnałów o zagrożeniu organizmu przegrzaniem. Model adaptacji do takich biegów, zachodzącej pod wpływem wyjątkowo silnego nacisku selekcyjnego, został opracowany z wykorzystaniem pracy Johna von Neumanna (zob. też architektura von Neumanna) na temat niezawodnych systemów z zawodnych elementów. Tworzenie „zapasowych” neuronów i równoległych połączeń między nimi miało doprowadzić do szybkiego wzrostu mózgu, który nie był skorelowany ze wzrostem sprawności myślenia. Twórcy modelu uważają, że ten proces był tylko preadaptacją do późniejszego rozwoju mózgu, np. powstania mowy (zob. ośrodek Wernickego, ośrodek Broki).
Pozycja człowieka w przyrodzie
Człowiek jest ssakiem łożyskowym z najlepiej wśród wszystkich zwierząt dostosowanym mózgiem do myślenia abstrakcyjnego. Człowiek jest przedstawicielem ssaków naczelnych (Primates), i ściślej – małp wąskonosych (Catarrhini). Człekokształtne naczelne wyróżnia brak ogona, większe rozmiary ciała i powiększony w stosunku do masy ciała mózg. Do człowiekowatych (Hominidae) zalicza się obecnie obok człowieka jego wymarłe formy oraz orangutany, goryle i szympansy.
Analogicznie jak przy wyjaśnianiu okoliczności powstania życia na Ziemi, pełne wyjaśnienie okoliczności towarzyszących powstaniu człowieka powinno ustalić kolejność powstawania charakteryzujących go cech taksonomicznych, za które – w dużym uproszczeniu – przyjmuje się najczęściej osiągnięcie dwunożnego chodu przy pionizacji postawy ciała oraz rozwój mózgowia, a zatem i inteligencji, rozumianej jako myślenie (również abstrakcyjne), pogłębienie zdolności uczenia się i rozwiązywania problemów. Nie dysponujemy jeszcze pełną teorią antropogenezy. Poniżej przedstawiono próbę podsumowania obecnego stanu wiedzy w tej dziedzinie.
Rys historyczny myśli ewolucyjnej

Podobieństwa pomiędzy ludźmi i niektórymi dużymi małpami człekokształtnymi dostrzegano od bardzo dawna. Malajczycy nazywali orangutana leśnym człowiekiem (orang-outang), Karol Linneusz zaklasyfikował do rodzaju Homo człowieka i szympansa, któremu nadał nazwę systematyczną Homo nocturnus (nocny człowiek, człowiek nocy), a rodzinę człowiekowatych (Hominidae) umieścił wśród małp człekokształtnych.
Kolejny etap rozwoju myśli ewolucyjnej rozpoczął się od odkrycia w 1829 (Engis, Belgia), w 1848 (Forbes’ Quarry, Gibraltar) i w 1856 roku (Neanderthal, Niemcy) kości istot podobnych do człowieka (neandertalczyk). Pojawiło się wówczas pojęcie człowieka jaskiniowego, które przetrwało do naszych czasów w kulturze masowej. Do czasów Darwina w świecie chrześcijańskim panowało powszechne przekonanie o wyjątkowej pozycji człowieka wśród żywych stworzeń (antropocentryzm). Opublikowana przez Darwina teoria ewolucji wywołała lawinę dyskusji pomiędzy naukowcami (Thomas Henry Huxley, Georges Cuvier, Charles Lyell, Richard Owen, Alfred R. Wallace i inni) oraz w wielu innych kręgach.
Karol Darwin zakładał, że poszukiwania skamieniałości wczesnych hominidów powinny być prowadzone w Afryce (Darwin, 1871). Wnioskował tak z analizy zasięgu naturalnego występowania gatunków najbliżej spokrewnionych z człowiekiem.
Poszukiwania prowadzone w różnych regionach świata – poza młodszymi genealogicznie okazami – nie dostarczały nowych danych o formach starszych od neandertalczyka. Począwszy od 1924 sytuacja zaczęła się zmieniać. Wskazywany przez Darwina kierunek poszukiwań wydawał się właściwy i kolejne lata przynosiły coraz to nowe odkrycia. Uwaga całego świata skoncentrowała się na afrykańskich stanowiskach paleoantropologicznych.

W miarę napływających doniesień z wyników badań nad znalezionymi skamieniałościami obraz drogi rozwojowej człowieka ulegał zmianie. Pojawiły się naukowe opisy odkrytych szczątków, pierwsze próby ich klasyfikowania (Australopithecus, Plesianthropus, Paranthropus, Zinjanthropus, Gigantopithecus, Oreopithecus bambolii, Meganthropus, Pithecanthropus, Palaeoanthropus, Cyphanthropus, Euranthropus, Protanthropus) oraz pierwsze próby rekonstrukcji ich wyglądu. Kolejne odkrycia dostarczały nowych danych, często zmieniając dotychczasowy obraz antropogenezy.
Filogeneza
Wczesna ewolucja naczelnych
Pierwsze łożyskowce (Eomaia) pojawiły się w kredzie z grupy wymarłych, żyworodnych ssaków, nazywanych Eupantotheria, Eomaia prawdopodobnie dała początek kilku grupom: Xenarthra, Afrotheria, Euarchontoglires, Laurasiatheria oraz Protoeutheria. Pierwsze euarchonty (Plesiadapis), w paleocenie dały początek pierwotnym małpiatkom, te dały początek linii ewolucyjnej, której pierwszym przedstawicielem był Eosimias, żyjący 45–40 mln lat temu. Do tej linii ewolucyjnej obecnie należą małpy szerokonose, makakokształtne oraz małpy człekokształtne.
Na podstawie analizy wyników badań molekularnych ustalono, że małpy szerokonose (Platyrrhini) oddzieliły się od wąskonosych ok. 35 mln lat temu Nie ma pewności, co do tego, gdzie wyewoluowały naczelne. Wielu badaczy skłania się ku Afryce, jednak prace archeologów na terenie Eurazji zdają się temu przeczyć. Niektórzy naukowcy wskazują nawet na Amerykę Północną.
Oligocen
Wszystkie naczelne wyższe, które nie należą do małp szerokonosych (Platyrrhini), zamieszkujących Nowy Świat, klasyfikowane są jako małpy wąskonose. Do najwcześniejszych wąskonosych zaliczamy m.in. rodzaje Propliopithecus i Parapithecus, których pozostałości odkryto w Fajum, niedaleko Kairu w pokładach z wczesnego oligocenu (35 – 31 mln lat temu). W tamtym czasie tereny stanowiska w Fajum porastał gęsty las tropikalny. Wszystkie naczelne z Fajum (prócz wymienionych dwóch występowały tam także przynajmniej dwa inne wąskonose oraz sześć bardziej prymitywnych naczelnych) żyły w górnych partiach lasu, do czego miały znakomicie przystosowane kończyny. Z prehominidów z Fajum rozwinęły się formy, takie jak Saadanius, żyjący 29–28 mln lat temu. W tamtym czasie z jednego z krewnych Saadaniusa wyewoluowały dwie główne linie rozwojowe wąskonosych: człekokształtne (Hominoidea) i zwierzokształtne (Cercopithecoidea), obecnie reprezentowane przez makakowate.
Rozwój człekokształtnych
Radiacja w miocenie
Do jednych z najwcześniejszych człekokształtnych zaliczany jest rodzaj Proconsul – małpa, żyjąca już ok. 27 mln lat temu, zaledwie 1–2 mln po rozdzieleniu się wąskonosych na człekokształtne i zwierzokształtne. Był on znacznie większy od pierwotnych form z Fajum, posiadał także bardziej rozwinięty mózg. Większość gatunków Proconsula zamieszkiwała wilgotny las tropikalny, choć przynajmniej jeden żył w bardziej suchym środowisku. Małpy te żyły przede wszystkim w Afryce Wschodniej. Około 20–15 mln lat temu nastąpiła gwałtowna radiacja człekokształtnych, związana z wyewoluowaniem z Proconsuli bardziej rozwiniętych form, do których zaliczamy gibony (Hylobatidae) i afropiteki (Afropithecoinea), a do tych zalicza się przede wszystkim zamieszkujące Afrykę Wschodnią Afropithecus i Kenyapithecus.

Późnomioceńskie (15–5 mln lat temu) naczelne określane są jako hominoidy. Charakteryzuje je:
- brak ogona
- zatrzymanie rozrostu kości miednicy
- budowa ramion i barków umożliwiająca zawieszanie się na gałęziach
- mózg nieznacznie większy niż u współczesnych małp (z uwzględnieniem proporcji ciała)
- rozwój wzroku
Driopiteki
Około 14 mln lat temu zapewne z prymitywniejszych, afrykańskich form wyewoluowała rodzina Dryopithecidae, której typowym przedstawicielem był driopitek, zamieszkujący tereny Europy i Chin. Charakteryzował się on grubszymi, potężniejszymi kośćmi w porównaniu z afropitekami i formami im pokrewnymi, jednak jego szczęka była drobniejsza, szkliwo zaś cieńsze, co utrudnia jednoznaczne umiejscowienie driopiteków na drzewie filogenetycznym człekokształtnych: nie jest pewne która grupa była bliżej spokrewniona z człowiekowatymi: czy driopiteki czy ich afrykańscy krewni, choć obecnie większość uczonych skłania się ku tym pierwszym. Driopiteki zapewne dały początek żyjącym w Europie człekokształtnym, takim jak Orepithecus bambolii czy Anoiapithecus, który jest jednym spośród kilku możliwych antenatów rodziny człowiekowate (Hominidae).
Ewolucja orangowatych
Orangowate (Ponginae), których obecnym przedstawicielem jest orangutan (Pongo) wyewoluowały najprawdopodobniej w Afryce około 13–12 mln lat temu z afropiteków, form im pokrewnych lub z driopiteków. Za najstarszy rodzaj uznawany jest Sivapithecus, którego skamieniałości odkryto w Kenii i Indiach. Siwapiteki żyły w okresie od ponad 12 do 8 mln lat i dały początek szeregowi pokrewnych form, jak Ouranopithecus żyjący w Grecji, Ankarapithecus, zamieszkujący Azję Mniejszą, czy Lufengpithecus z Chin. Ich bezpośrednim potomkami były także żyjące obecnie dwa gatunki orangutanów (Pongo): borneański (Pongo pygmaeus) oraz sumatrzański (Pongo abelii). Około 1 mln lat temu pojawił się rodzaj Gigantopithecus, którego przedstawiciele dorastali do 3 m i osiągali masę do 540 kg. Ta występująca w Indiach, Chinach i Wietnamie małpa jest uważana za największego naczelnego w dziejach. Wymarła stosunkowo niedawno – 300 tysięcy lat temu.
Wczesne hominidy
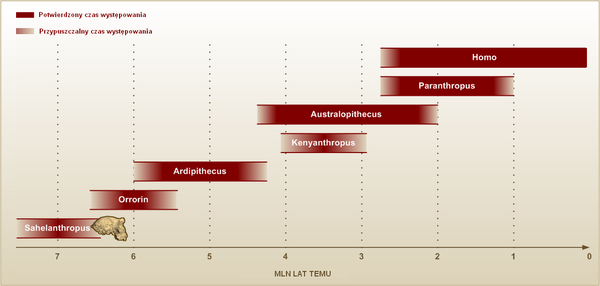
Mimo że termin „hominidy” oznacza przedstawicieli rodziny człowiekowatych, nieoficjalnie przyjęło się, iż hominidami są jedynie naczelne, należące do linii rozwojowej człowieka, charakteryzujące się dwunożną postawą ciała, znacznym wzrostem mózgu w stosunku do proporcji ciała, zmianą w uzębieniu i muskulaturze, naziemnym tryb życia, długotrwałym rozwojem osobniczym. Pierwsze hominidy pojawiły się zapewne w okresie między od 14–10 do 7–6 mln lat temu, kiedy najpierw goryle (11–8 mln lat temu), a potem szympansy (7–5) oddzieliły się od linii rozwojowej człowieka. Okres ten jest bardzo słabo poznany, skamieniałości są nieliczne, a te, które udało się odnaleźć trudno porównać z kośćmi dzisiejszych małp i ludzi.
Ostatnio przyjmuje się coraz szerzej używanie nazwy homininy zamiast hominidy, odkąd zaproponowano wydzielenie nowej, osobnej jednostki taksonomicznej – podplemienia Hominina.
Wspólni przodkowie małp człekokształtnych i ludzi
Ostatnie odkrycia człowiekowatych, pochodzących z południowej Europy zdają się podważać teorię, zakładającą, iż pierwsze hominidy pojawiły się w Afryce. Małpy człekokształtne z Europy były zapewne potomkami driopiteków, które mogły się tam dostać poprzez Półwysep Arabski lub pomost lądowy pomiędzy dzisiejszym Marokiem i Półwyspem Iberyjskim. Wśród południowoeuropejskich człekokształtnych z późnego miocenu były takie gatunki jak Pierolapithecus catalaunicus z Katalonii (Hiszpania), sprzed 13 mln lat. Inne, młodsze znaleziska pochodzą jednak z Afryki Wschodniej, jak skamieniałości Chororapithecus abyssinicus (ok. 8 mln lat temu), uważany przez wielu uczonych za przodka współczesnych goryli, Samburupithecus kiptalami (9,5 mln), Nakalipithecus nakayamai (10 mln). Istnieje hipoteza, że najstarsi przodkowie hominidów wyewoluowały z dryopiteków w Europie i na Bliskim Wschodzie, skąd ponownie przybyli do Afryki. Na podstawie nielicznych znalezisk sprzed 7 mln lat temu nie można powiedzieć wiele na temat wspólnego przodka goryli, szympansów i ludzi. Pewnych informacji dostarcza pochodząca sprzed 7 mln lat czaszka Sahelanthropus tchadensis, który miał bardzo duże wały nadoczodołowe, małą puszkę mózgową, średnio wysuniętą do przodu trzewioczaszkę i silnie do tyłu mózgoczaszkę.
Sahelanthropus tchadensis

W latach 2001 Michel Brunet opisał nowy gatunek hominida – Sahelanthropus tchadensis, którego czaszkę liczącą ok. 7 mln lat odkryto w Czadzie. Brak szczątków sahelantropa, prócz kości czaszki uniemożliwia stwierdzenie, czy poruszał się on w pozycji dwunożnej, czy też jego chód przypominał sposób poruszania się dzisiejszych małp człekokształtnych. Odkrywca sahelantropa – Michel Brunet uważa, że cechy kłów bardziej przypominają cechy człowieka, niż goryla czy szympansa. Mimo tego jednak naczelny ten posiada wiele cech, upodobniających go do małp człekokształtnych: małą puszkę mózgową, szeroko rozstawione oczodoły. Brunet uznaje to za dowód na bliskie pokrewieństwo z ostatnim wspólnym przodkiem szympansa, goryla i człowieka i proponuje uznać go za protoplastę wszystkich późniejszych hominidów, w tym orrorina i ardipiteka. Z opinią Bruneta nie zgadza się wielu naukowców, którzy sceptycznie oceniają jego twierdzenie o hominidalnym charakterze kłów sahelantropa. Uważają oni, że masywne wały nadoczodołowe, mała puszka mózgowa i inne cechy upodobniają go do małp. Odnalezienie szczątków sahelantropa w Czadzie zadało ostateczny cios teorii, według której przodkowie szympansów i ludzi rozdzielili się, gdy kontynent afrykański pękł na dwie części. Zgodnie z tą hipotezą hominidy ze wschodniej części (dzisiejsza Kenia, Etiopia, Uganda, Somalia, Tanzania) dały początek australopitekom i człowiekowi, te z zachodniej części (Dolina Kongo, Czad) natomiast – szympansom.
Orrorin tugenensis
Wśród innych hominidów z tamtego okresu znamy Orrorin tugenensis, którego szczątki odkryli po raz pierwszy w 2001 roku paleontolodzy Martin Pickford i Brigitte Senut z Musèum Natrional d'Historie Naturelle w Paryżu. Badacze ci odnaleźli fragmenty żuchwy, pojedyncze zęby, kości palców, kości ramienne i fragmenty kości udowych, pochodzących sprzed ok. 6 mln lat. Według odkrywców orrorina, naczelny ten jest niewątpliwie przedstawicielem człowiekowatych, skamieniałości potwierdzają zaś, iż poruszał się na dwóch nogach. Utrzymanie wyprostowanej pozycji ułatwiał znacznie przyczepiony do dłuższej niż u wcześniejszych naczelnych szyjki uda, mięsień zasłaniacz zewnętrzny, obracający udo na zewnątrz. Mimo dwunożnego chodu orrorin miał także wiele cech wczesnych naczelnych: duże, ostro zakończone kły oraz przystosowane do wspinania się po drzewach kończyny górne i palce. Jego odkrywcy: Martin Pickford i Brigitte Senut uważają, że jest on bardziej podobny i bliżej spokrewniony z Homo, aniżeli większość australopiteków (wśród nich jest słynna Lucy) i parantropy. Zaproponowali utworzenie osobnego rodzaju: Praeanthropus, do którego zaliczyli prócz orrorina część skamieniałości obecnie klasyfikowanych do gatunków Australopithecus afarensis i Australopithecus anamensis. Według nich wszystkie inne australopiteki i wywodzące się od nich parantropy są niezależną i bezpotomną linią rozwojową, ardipiteki natomiast dały początek szympansowi. Ustalenia Pickforda i Senut nie przekonują wielu uczonych, którzy uważają, iż orrorin, mimo że mógł chodzić na dwóch nogach, spędzał wiele czasu na drzewach i bardziej przypominał małpy człekokształtne, aniżeli człowieka. Badacze ci jednak nie negują pokrewieństwa tegoż hominida z nami, a uznają go za znacznie dalszego krewnego lub przodka, niż sądzili to Pickford i Senut.
Ardipithecus kadabba
Kilka miesięcy po odkryciu i opisaniu Orrorin tugensis Yohannes Haile-Selas, doktorant z Berkeley opublikował informację o odkryciu skamieniałości hominida w regionie środkowego Anuaszu w Etiopii, sprzed ok. 5,2 – 5,8 mln lat. Szczątki zaklasyfikowano do znanego od lat 90. XX wieku gatunku Ardipithecus ramidus, jednak wydzielono osobny podgatunek, nazwany A. r. kadabba. W późniejszym czasie skorygowano ten podział i odtąd skamieniałości zaliczano do osobnego gatunku Ardipithecus kadabba. Badający skamieniałości nogi ardipiteka C. Owen Lovejoy z Kent State University stwierdził, że hominid ten chodził bez wątpienia podobnie jak człowiek. Mimo tego wielu badaczy podało w wątpliwość dwunożny chód ardipiteka, uznając, że jego stopa przypomina bardziej stopę szympansią, aniżeli ludzką.
Ardipithecus ramidus
Nieco późniejszym hominidem niż Ardipithecus kadabba był Ardipithecus ramidus, którego szczątki pochodzące sprzed ok. 4,4 mln lat temu odkryto w 1995 roku. Posiadał on cechy charakterystyczne dla małp, ale również pewne cechy charakterystyczne już dla ludzi. Na przykład kształt i położenie otworu potylicznego wielkiego (foramen magnum) w podstawie czaszki, przez który rdzeń kręgowy łączy się z mózgiem, przypomina kształt tego otworu, jaki mają ludzie. Cecha ta – obok kilku innych – świadczy o opanowaniu umiejętności poruszania się na dwóch nogach (dwunożności).
Hipoteza Owena C. Lovejoya dotycząca genezy monogamii i dwunożności u homininów
Antropolog z Uniwersytetu Stanowego w Kent w stanie Ohio prof. Owen Lovejoy pokusił się o skonstruowanie modelu mającego wyjaśnić pochodzenie specyficznych cech ardipiteków. Dużą rolę odgrywa w tym modelu unikalna próba rekonstrukcji ich życia seksualnego i rozrodczego. Obszerną prezentację tego modelu w polskim piśmiennictwie można znaleźć w książce Jerzego A. Kowalskiego „Homo eroticus” z 2011 roku. Punktem wyjścia rekonstrukcji jest zaskakująco niski poziom dymorfizmu płciowego, szczególnie w zakresie wielkości kłów. Kły służą zwierzętom nie tylko do walki, ale także do zmagań o ustalenie statusu w obrębie stada. Ostra rywalizacja o samice sprzyja więc powiększaniu się wielkości kłów u samców i prowadzi do zwiększania ich dymorfizmu. Skoro u ardipiteków został zredukowany dymorfizm kłów, to nasuwa się wniosek, że stał za tym bardzo niski stopień rywalizacji pomiędzy samcami. Niski poziom rywalizacji wskazuje zwykle na istnienie bardziej wyrównanego dostępu samców do partnerek. Lovejoy wysnuwa więc wniosek, że w grupach ardipiteków podstawę systemu kojarzenia stanowiły monogamiczne pary.
Hipotezę Lovejoya polski autor nazywa hipotezą „samców-zaopatrzeniowców”. Wedle niej męskie osobniki na szeroką skalę zaopatrywały wybrane przez siebie partnerki w żywność. Było to konieczne, ponieważ rozrastający się u ardipiteków mózg wymagał dłuższego okresu dojrzewania, a zatem dłuższej ciąży i dłuższego dzieciństwa, co czyniło matki mniej sprawnymi i mniej samodzielnymi. Potrzebna im była pomoc żywnościowa, zwłaszcza że z powodów klimatycznych kurczyły się w owym czasie zasoby pokarmowe. Samice preferowały jako swoich partnerów nieagresywne, opiekuńcze samce, one bowiem najbardziej nadawały się na „zaopatrzeniowców”. Samce agresywne i dominujące były odsuwane przez samice na dalszy plan, wskutek czego obniżał się poziom rywalizacji i antagonizm pomiędzy samcami. Podstawą społeczności ardipiteków stały się zatem monogamiczne pary. Drastycznym, dobrze widocznym, niejako ubocznym skutkiem działalności aprowizacyjnej samców stała się przy tej okazji dwunożność homininów. Tylko bowiem w wyprostowanej pozycji, z uwolnioną parą rąk możliwe było przenoszenie większych ilości pożywienia na dłuższy dystans. Jak widać, w poręczny sposób koncepcja ta za jednym zamachem próbuje wytłumaczyć zarówno przyjęcie przez ardipiteki monogamii, jak i unikatowe, specyficzne dla homininów wyprostowanie postawy ciała.
Dwunożność
Za jedną z najważniejszych cech odróżniających hominidy od innych małp, jak goryle, czy szympansy jest dwunożność. Dawniej większość naukowców uważała, iż dwunożny tryb życia nasi przodkowie przyjęli, gdy zmuszeni zostali do życia na sawannach. Gdyby jednak okazało się, że Orrorin tugenensis i Ardipithecus kadabba poruszały się w pozycji wyprostowanej teoria ta zostałaby obalona, oba te gatunki zamieszkiwały bowiem wilgotne, tropikalne lasy, gdzie razem z nimi żyły leśne małpy, słonie i inne zwierzęta. Giday Wolde-Gabriel wraz ze współpracownikami z Los Alamos National Laboratory, badający skład chemiczny gleby i szczątki zwierzęce znalezione na stanowisku, na którym odkryto szczątki Ardipithecus kadabba, dowiedli, że wczesne hominidy zamieszkiwały tamte tereny do przynajmniej 4,4 mln lat temu.
Australopiteki
Żyły od ok. 4,2 do ok. 1 mln lat temu (pliocen i wczesny plejstocen), początkowo w Afryce Wschodniej, na terytorium Wielkich Rowów Afrykańskich – tam naukowcy doszukują się kolebki ludzkości – później także w Afryce Południowej i Środkowej. Od swoich poprzedników różniły się głównie dwiema cechami: większym o około 20% mózgiem oraz mocniejszym uzębieniem. Charakteryzowały się masywną szczęką, mocnym uzębieniem, ale już krótkimi kłami – umożliwiało to poziomy ruch trący i świadczy o dużym udziale twardego pokarmu roślinnego w ich pożywieniu (orzechy, kłącze, bulwy, nasiona). Podobnie jak wcześniejsze ardipiteki miały stopy o nieprzeciwstawnym paluchu (jak u człowieka), były przystosowane do chodzenia w pozycji pionowej, czyli do dwunożności, o czym świadczyć może m.in. budowa podstawy czaszki i położenie foramen magnum w odniesieniu do płaszczyzny przechodzącej przez górny i dolny brzeg oczodołu (tzw. kąt alfa), struktura uzębienia, czy budowa pasa biodrowego.
Koncepcje dotyczące wytworzenia się nagiej skóry
Wśród naczelnych tylko człowiek pozbył się owłosienia prawie całego ciała. Naga skóra jest też zupełną rzadkością u całej gromady ssaków. Naukowcy od dawna zastanawiali się nad sensem i mechanizmem zrzucenia owłosienia ciała przez ludzi. Karol Darwin uważał nagość naszej skóry za cechę wykształconą wskutek doboru płciowego, czyli mającą znaczenie dla systemu kojarzenia, a nie bezpośrednio dla przeżywalności gatunku.
Najbardziej do dzisiaj wpływową koncepcją wyjaśniającą biologiczną funkcję nagiej skóry jest idea człowieka jako pewnego rodzaju „nagiej małpy”, którą spopularyzował Desmond Morris. Wedle tej koncepcji głównym czynnikiem sprawczym zaniku owłosienia ciała u ludzi miałaby być konieczność chłodzenia organizmu przegrzewającego się podczas biegu towarzyszącego polowaniom. Ponieważ polowaniem zajmuje się wiele gatunków drapieżnych zwierząt, a żaden z nich nie zrzucił futra tak jak my, dodaje się zwykle zastrzeżenie, że chodzi o specyficzną dla ludzi metodę polowania, a mianowicie ściganie ofiary aż do jej całkowitego zmęczenia i dopadanie jej, gdy już nie mogła dalej uciekać. Antropolog Richard Wrangham w jednej z ostatnich swoich publikacji nieznacznie uzupełnia tę koncepcję o dodatkowy czynnik w postaci zastosowania ogniska jako źródła ciepła, które pozwalało nagim osobnikom ogrzewać się w czasie chłodniejszych nocy. Na ogół uważa się przy tym, że pierwszym gatunkiem, który utracił owłosienie całego ciała, był Homo erectus.
Najnowsze porównawcze badania genetyczne nad pasożytami człowiekowatych zdają się jednak wskazywać, że już australopiteki około 3,3 mln lat temu posiadały bezwłosą, nagą skórę na całym prawie ciele. Jerzy A. Kowalski zaproponował ostatnio wyjaśnienie, wedle którego nagość skóry – jak chciał K. Darwin – jest cechą płciową. Jej zaczątki miały się pojawić już u samic ardipiteków jako uboczny skutek przyjęcia wyprostowanej postawy ciała, przy której dawne, charakterystyczne np. dla szympansów obrzmienia genitalne samic musiały z przyczyn czysto mechanicznych zaniknąć. W tym miejscu pozostał płat nagiej skóry, tzw. skóry płciowej, który wskutek doboru płciowego rozszerzył się na całe ciało.
Inną, konkurencyjną teorią jest tzw. hipoteza wodnej małpy, która nie zdobyła sobie jednak szerszej akceptacji.
Australopithecus anamensis
Większość naukowców zgadza się ze stwierdzeniem, że bezpośrednimi potomkami Ardipithecus ramidus były australopiteki – hominidy, które pojawiły się ok. 4,2 mln lat temu. Najstarszym znanym gatunkiem należącym do nich był Australopithecus anamensis, żyjący w Kenii. Była to bez wątpienia istota, poruszająca się na dwóch nogach, o czym świadczy budowa kończyn dolnych, posiadała szkliwo znacznie grubsze niż u swoich przodków, co pozwala sądzić, że żywiła się twardym pokarmem, jednak budowa szczęki i niektórych części czaszki upodobniała ją do małp człekokształtnych. W środowisku, w którym żył tenże hominid przeważały, zamieszkiwane przez antylopy oraz bawolce sawanna, choć nie brakowało tam wilgotnych, gęstych lasów.
Australopithecus afarensis
Nieco młodszym, niż Australopithecus anamensis gatunkiem był, żyjący od ok. 3,6 do 2,9 mln lat temu Australopithecus afarensis, który zamieszkiwał tereny wschodniej Afryki, głównie Etiopii i Tanzanii. Jest to chyba jeden z najlepiej zbadanych australopiteków, głównie dzięki znalezieniu prawie kompletnego szkieletu, nazwanego Lucy. Wokół gatunku tego istnieje wiele kontrowersji, np. czy znaleziska etiopskie i tanzańskie należą do tego samego gatunku lub czy wiek okazów został poprawnie określony. Hominid ten żył prawdopodobnie w środowisku, gdzie przeważały sawanny, mało czasu spędzał więc prawdopodobnie na drzewach, choć zapewne przebywał tam w czasie snu, aby nie być narażonym na ataki drapieżników. Charakteryzował go mały mózg, nieco większy od mózgu szympansa, mocne szczęki, pokryte grubym szkliwem zęby (co pomagało w rozcieraniu twardego pokarmu) oraz wyraźny dymorfizm płciowy: samce były znacznie większe od samic. Australopiteki żyły w małych grupach rodzinnych, w których, jak się zakłada, istniała hierarchia, a nawet podział pracy w zależności od płci. Żywiły się na ogół owocami, roślinami, owadami, małymi zwierzętami, niekiedy także padliną. Porozumiewały się zapewne różnego rodzaju okrzykami i gestami.
Kenyanthropus platyops
W 1999 roku nad Jeziorem Turukana w Kenii odkryto szczątki hominida, sprzed 3,5 – 3,2 mln lat temu, nazwanego Kenyanthropus platyops. Kenyanthropus charakteryzował się małą puszką mózgową, jednak wysokie kości policzkowe upodobniały go do rodzaju Homo. Poruszał się zapewne w postaci wyprostowanej, zamieszkiwał rozległe sawanny, które dzielił z Australopithecus afarensis'. Toczą się liczne spory o pokrewieństwo tegoż hominida z innymi przedstawicielami rodziny człowiekowatych. Wielu uczonych uważa, że był on jedynie lokalną odmianą Australopithecus afarensis, inni przypuszczają natomiast, że należy on do odrębnej linii ewolucyjnej, która mogła dać początek rodzajowi Homo, a konkretnie Homo rudolfensis.
Australopithecus bahrelghazali
W 1993 roku francuski badacz Michel Brunet odkrył na terenie Czadu szczątki australopiteka, pochodzące sprzed 3 – 3,5 mln lat, nad których klasyfikacją trwają spory: jedni uważają, że należą one do odrębnego gatunku Australopithecus bahrelghazali, inni sądzą, iż jest to odmiana lokalna Australopithecus afarensis. Odkrycie skamieniałości australopiteka ponad 2,5 tysiąca kilometrów od Wielkiego Rowu Tektonicznego, który, jak wcześniej zakładano, stanowił granicę terytoriów, zasiedlanych przez te hominidy doprowadziło do rewizji ówczesnych hipotez i ukazało, iż zamieszkiwały one znacznie rozleglejsze obszary Afryki. Australopithecus bahrelghazali zamieszkiwał zarówno wilgotne lasy tropikalne, jak i sawanny i łąki. Anatomia jego żuchwy przypominała budowę żuchwy Australopithecus afarnesis, jednak posiadające trzy korzenie zęby trzonowe był podobne do zębów hominidów z rodzaju Paranthropus, który wyewoluował z australopiteków ok. 2,5 mln lat temu.
Australopithecus africanus
Podczas gdy Australopithecus afarensis zamieszkiwał tereny Afryki Wschodniej, na południu w okresie od ok. 3 do 2 mln lat temu żył inny gatunek hominida – Australopithecus africanus. Typowy jego męski przedstawiciel mierzył ok. 1,3 m, ważył 30 – 40 kg, żeński zaś mierzył ok. 1,15 m i ważył ok. 30 kg. Australopitek miał niezbyt dużą puszkę mózgową. Głównym jego pożywieniem były korzenie, bulwy, nasiona, w mniejszych ilościach owoce, liście, także mięso. O środowisku, w którym żył wiadomo niewiele. Pierwotnie zapewne tereny Afryki Południowej były bardziej wilgotne niż dzisiaj, jednak z biegiem czasu klimat osuszał się, co prowadziło do zanikania buszów i rozprzestrzeniania się sawann.
Australopithecus sediba
W 2010 roku opisano nowy gatunek hominida – Australopithecus sediba, którego szczątki odkryto w położonej w Afryce Południowej jaskini Malapa, około 40 km od Johannesburga, który, jak sądzi większość naukowców, wyewoluował z Australopithecus afarensis, blisko z nim spokrewnionego Australopithecus garhi lub od Australopithecus africanus. Hominid ten łączy w sobie zarówno cechy prymitywne, np. mały mózg, drobną posturę, czy długie ramiona, z cechami nowoczesnymi, jak płaską twarzą, długimi nogami i silną miednicą, przystosowaną do sprawnego chodzenia. Wielu uczonych dostrzega w szkielecie Australopitecus sediba podobieństwo do pierwszych przedstawicieli Homo, dawniej uważano tenże gatunek za bezpośredniego przodka praludzi, jednak po opisaniu żyjącego równolegle z nim, bardziej nowoczesnego Homo gautengensis uznaje się go za boczną, ślepą gałąź na drzewie genealogicznym hominidów.
Paranthropus
Australopithecus afarensis dał prawdopodobnie początek zarówno młodszym formom australopiteków, jak i niezależnemu, bezpotomnemu rodzajowi, nazwanemu Paranthropus, do którego zaliczamy trzy gatunki: P. aethiopicus oraz wywodzące się od niego P. robustus, który zachował wiele cech pierwotnych, i P. boisei, który uległ znacznej specjalizacji. Początkowo parantropy wyobrażano sobie jako masywne, potężne hominidy, w rzeczywistości jednak miały one jedynie silne, duże, mocno wysunięte szczęki, przystosowane do rozcierania twardych pokarmów, np. orzechów, a posturą ciała nie różniły się od australopiteków. Ostatnie z parantropów żyły ok. 1 mln lat temu, równolegle z pierwszymi przedstawicielami rodzaju Homo. Istnieje wiele teorii na temat przyczyn ich wymarcia: jedna z nich mówi, że hominidy te przegrały rywalizację z ludźmi, którzy byli znacznie bardziej wszechstronni i, co najważniejsze, posiadali umiejętność wytwarzania kamiennych narzędzi.
Ewolucja rodzaju Homo
Zachowania narzędziowe
Pierwsi ludzie (łac. Homo) wyewoluowali zapewne z któregoś z gatunków australopiteków ponad 2,5 mln lat temu. Mieli oni, w porównaniu ze swoimi przodkami, większe mózgi, byli nieco wyżsi oraz, co najważniejsze, posługiwali się kamiennymi narzędziami otoczakowymi. Ich wynalezienie około 2,5 mln lat temu przypisuje się wprawdzie już australopitekom, jednak pierwsi ludzie udoskonalili je na tyle, że zasłużyły na miano kultury olduwajskiej. Wynalezienie narzędzi otoczakowych umożliwiło im obronę przed dzikimi zwierzętami, a także pozwoliło udoskonalić metody polowania. Odtąd mięso stawało się coraz ważniejszym elementem diety. Niektórzy naukowcy przypuszczają, że miało to duże znaczenie dla poprawy bilansu energetycznego organizmu człowieka, pozwalającego na wykształcenie się coraz większego mózgu oraz mogło się przyczynić do rozwoju coraz bardziej złożonych form społecznych.
Najstarsi przedstawiciele Homo
Najstarsi ludzie pojawili się ponad 2,5 mln lat temu, prawdopodobnie w Południowej Afryce, gdzie żył najstarszy znany gatunek Homo gautengensis. Zamieszkiwany przez niego obszar sugeruje, iż mógł wywodzić się od Australopithecus africanus. Gautengensis był niewysoki (mierzył ok. 90 cm), miał jednak większy mózg od swoich przodków, zapewne posługiwał się kamiennymi narzędziami. Wśród nieco późniejszych praludzi, które zamieszkiwały tereny Afryki Wschodniej znajdują się Homo habilis (pojawił się ok. 2,3 mln lat temu) i Homo rudolfensis (występował od 2,4 mln lat temu), których wzajemne pokrewieństwo, a także relacje z Homo gautengensis pozostają nieznane. Wyżej wymieniony Homo habilis posiadał dość duży mózg (ok. 750 cm³), jednak jego wysunięta do przodu twarzoczaszka i silne wały nadoczodołowe upodobniały go do wczesnych hominidów. Drugi, wschodnioafrykański praczłowiek – Homo rudolfensis, mimo mniejszej puszki mózgowej (ponad 500 cm³), miał twarzoczaszkę bardziej przypominającą twarz człowieka współczesnego.
Pochodzenie rodzaju Homo
Homo ergaster
Około 1,9–1,8 mln lat temu z któregoś z wczesnych gatunków Homo w Afryce Wschodniej wyewoluował znacznie bardziej rozwinięty, dużo wyższy od swoich przodków (był to rodzaj ochrony przed upałem: większa powierzchnia ciała umożliwiała wydalanie więcej potu, który ochładzał ciało) Homo ergaster. Podstawową cechą wyróżniającą go od wcześniejszych hominidów jest znacznie bardziej rozwinięty mózg, a szczególnie płat czołowy, odpowiadający za posługiwanie się mową, panowanie nad emocjami, myślenie symboliczne. Zwiększona inteligencja umożliwiła wytwarzanie (od ok. 1,5 mln lat) nowych rodzajów narzędzi: czoperów i odłupków w przemyśle olduwajskim, a potem rozłupców i pięściaków w przemyśle aszelskim. Ponadto Homo ergaster posiadał znacznie mniejsze kły, co świadczyło o zaniku dymorfizmu płciowego – cechy występującej u małp i wczesnych hominidów. Wraz z powolnym zanikiem dymorfizmu i zwiększeniem się zdolności umysłowych ludzie zaczęli tworzyć coraz bardziej zorganizowane grupy, które dawały im ogromną przewagę nad innymi hominidami, które z czasem zostały przez nich wyparte.
Zasiedlenie Azji: 1,9–1,7 mln lat temu
Pierwsze próby ekspansji poza Afrykę podjął człowiek prawdopodobnie około 2–1,9 mln lat temu, czego dowodem są chociażby pochodzące sprzed 1,8 mln lat szczątki hominida, odkryte w 1991 roku w gruzińskim miasteczku Dmanisi. Odkrycie to wstrząsnęło światem nauki, bowiem dotychczas uważano, że jako pierwszy Afrykę opuścił około 1 mln lat temu Homo erectus. Objętość puszek mózgowych dwóch odkrytych czaszek wynosiła odpowiedni 770 i 650 cm³, a zatem była znacznie mniejsza, niż u Homo erectus, do którego pierwotnie zaliczono człowieka z Dmanisi. Poza tym anatomia czaszki przypominała budowę czaszki Homo habilis. Odkrycie szczątków Homo georgicus, bo tak właśnie zaczęto nazywać tegoż hominida pokazało, że do podjęcia ekspansji na północ nie był potrzebny duży mózg. Trasa ekspansji pierwotnych ludzi, którzy ok. 1,9–1,7 mln lat temu skolonizowali Azję biegła prawdopodobnie ze Wschodniej Afryki, przez Bliski Wschód, Indie, Półwysep Malajski do Indonezji i na tereny Chin. Niewykluczone, że właśnie ci pierwotni ludzie dali początek hominidowi, nazwanemu nieoficjalnie mianem Meganthropus, którego szczątki znaleziono na Jawie.
Homo erectus
Ponad milion lat temu Afrykę opuściła druga fala Homo ergaster. Liczne grupy ludzi pierwotnych zaczęły zasiedlać tereny Bliskiego Wschodu, skąd skolonizowały tereny Azji Środkowej, Indii i Dalekiego Wschodu. Azjatycka populacja przekształciła się z czasem w nowy gatunek – Homo erectus, nazywany dawniej Pithecanthropus erectus. Ludzie ci zasiedlili także Jawę, gdzie żył już prawdopodobnie inny gatunek hominida, zwany megantropem i wywodzący się być może od pierwotnych przedstawicieli Homo, którzy ok. 1,9–1,7 mln lat temu zasiedlili Azję. Homo erectus z pewnością posiadał już umiejętność panowania nad ogniem, jednak nie wiadomo, czy potrafił go rozniecać. Nie bał już się płomieni, a ogrzewał się ich ciepłem, piekł mięso i odstraszał nimi dzikie zwierzęta. Zamieszkiwał jaskinie lub tymczasowe obozowiska. Najdłużej Homo erectus przetrwał na Dalekim Wschodzie i na Jawie. Ziemie te stanowiły rodzaj odizolowanej wyspy, na której ewolucja biegła niezależnie od rozwoju hominidów afrykańskich. Dowodem tego jest fakt, iż nigdy na tamte tereny nie dotarły narzędzia kultury aszelskiej, która ogarnęła Afrykę. Ten rodzaj człowieka jest już bardzo zbliżony do człowieka rozumnego. Podobny był jego wzrost, mózg także był porównywalnych rozmiarów (800-1250 cm³). Do jego najbardziej charakterystycznych cech można zaliczyć grubsze kości, silnie zaznaczone łuki brwiowe oraz duże zęby.
Zasiedlenie Europy
Mimo oddzielenia się Homo erectus ewolucja Homo ergaster w Afryce trwała nadal. Można przypuszczać, że ok. 1 mln – 800 tys. lat temu tamtejsi ludzie zaczęli przechodzić przez istniejące wówczas połączenia lądowe do południowej Europy, gdzie znaleziono skamieniałości, zaliczane do gatunków Homo antecessor (Hiszpania) i Homo capranensis (Włochy). Jednak pierwszej udanej próby kolonizacji Europy dokonał prawdopodobnie dopiero Homo heidelbergensis, który 600 tysięcy lat temu pojawił się w Afryce. Miał on mózg nieco mniejszy od naszego, był wysoki, posługiwał się nowoczesnymi jak na tamte czasy narzędziami. Być może był pierwszym gatunkiem, który posługiwał się prymitywną mową oraz grzebał zmarłych. Homo heidelbergensis szybko opanował większość Zachodniej, Południowej i Środkowej Europy, wyruszył także na Bliski Wschód i do Azji, gdzie napotkał populację Homo erectus. Nie wiadomo jakie relacje panowały między tymi oboma gatunkami.
Neandertalczyk
Homo sapiens
Od stada do cywilizacji
Pierwotny Homo sapiens zajmował się głównie myślistwem. Prowadził koczowniczy tryb życia. Polowania dostarczały mu wysokokalorycznego mięsa, skór, którymi się okrywał oraz kości wykorzystywanych do wytwarzania coraz wydajniejszych narzędzi. W poszukiwaniu zwierzyny i nowych terenów łownych zaludniał nowe obszary.
Religia i kultura
Najstarsze ślady religii pojawiły się 100 tys. lat temu w postaci pochówków (wiara w życie pośmiertne) i rysunków naskalnych.
Około 40 tys. lat p.n.e. pojawiły się w Indiach pierwsze zabytki sztuki prehistorycznej w postaci obiektów malarstwa jaskiniowego. W toku rozwoju tej sztuki pojawiły się następnie ryty naskalne (petroglify) oraz formy rzeźbiarskie, w szczególności mała rzeźba mobilna (figurki).
Rolnictwo i hodowla
Wędrówki zwierząt poszukujących źródeł wody i żywności przyczyniły się prawdopodobnie do celowej ‘opieki’ nad szczególnie przydatnymi dla człowieka gatunkami. Już ok. 9000 – 8000 p.n.e. doszło do udomowienia pierwszych zwierząt (pies, owca, koza). Prawie 10 000 lat p.n.e. człowiek zasiał ziarna dziko rosnących roślin, a około 8 000, a w niektórych regionach do 3000 lat p.n.e. rozpoczął celową uprawę roli (zobacz: rolnictwo). Przyczyniło się to do zmiany trybu życia na osiadły. Zaczęły powstawać pierwsze osady, a ok. 6,8 tys. lat temu – pierwsze miasta.
W ten sposób archaiczny Homo sapiens przez 250 tys. lat opanował cały wolny od lodu świat i jako jedyny z rodzaju Homo nabył umiejętności pozwalające mu przetrwać do czasów, które są początkiem cywilizacji.
Zobacz też
Uwagi
Bibliografia
Polskojęzyczna literatura
- Jerzy Adam Kowalski, Homo eroticus, Seria: Eros i logos, Wydawnictwo IBS, Opole 2011
- Christopher Stringer, Robin McKie, Afrykański exodus. Pochodzenie człowieka współczesnego, Wydawnictwo Prószyński i S-ka, Warszawa 2003
- Steve Olson Mapowanie historii ludzkości – Przeszłość ukryta w naszych genach, 2003
- Roger Lewin Wprowadzenie do ewolucji człowieka, 2002
- Henryk Szarski Historia zwierząt kręgowych, PWN, 1998
- Jerzy Dzik, Wykłady
Polskojęzyczne artykuły
- Kate Wong Nasz najstarszy przodek?, Świat Nauki Wydanie Specjalne, Numer 3 2003
- Meave Leakey, Alan Walker Najstarsze afrykańskie hominidy, Świat Nauki Wydanie Specjalne, Numer 3 2003
- Ian Tattersall, Nie zawsze byliśmy sami, Świat Nauki Wydanie Specjalne, Numer 4/2000
- Kate Wong, Zagadkowi neandertalczycy, Świat Nauki Wydanie Specjalne, Numer 3 2003
- Ian Tattersall, Pożegnanie z Afryką, Świat Nauki Wydanie Specjalne, Numer 3 2003
- Alan G. Thorne, Milford H. Wolpoff, Policentryczna ewolucja człowieka, Świat Nauki Wydanie Specjalne, Numer 6 1992
- Kate Wong, Powikłany rodowód, Świat Nauki Wydanie Specjalne, Numer 3 2003
- Rebecca L. Cann, Allan C. Wilson, Afrykański rodowód ludzkości, Świat Nauki Wydanie Specjalne, Numer 6 1992
- Kate Wong, Pionier z Gruzji, Świat Nauki, Numer 1 2004
- Kate Wong, Pouczająca tragedia
- Rozmowa z João Zilhão, Czy neandertalczycy myśleli tak jak my?, Świat Nauki, Numer 7 2010.
Literatura i artykuły anglojęzyczne
- G.C. Conroy Reconstructing Human Origins: a modern synthesis New York: W. W. Norton, 2005
- Yohannes Haile-Selassie, Late Miocene Hominids from the Middle Awash, Ethiopia, Nature, tom 412, s. 178–181, 12 VII 2001
- Ian Tattersall, Jeffre H. Schwartz, Extnict Humans, Westview Press, 2001
Linki zewnętrzne
- W języku angielskim
- Paleoanthropology
- Discovery of Early Hominids. anthro.palomar.edu. [zarchiwizowane z tego adresu (2014-12-14)].
- Paleoanthropology in the 1990's
- The Evidence for Human Evolution
- Exploring the Creation/Evolution Controversy
- W języku polskim
- Antropogeneza. ewolucja.org. [zarchiwizowane z tego adresu (2016-04-04)].
| Homininae | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Australopithecine |
|
||||||||
| Człowiek |
|
||||||||
| Tematy powiązane | |||||||||
![]()


![]()
| Procesy ewolucyjne | |
|---|---|
| Nauki związane z ewolucją | |
| Rodzaje specjacji | |
| Rodzaje doboru naturalnego | |
| Ewolucja molekularna | |
| Ewolucja poszczególnych taksonów | |
| Pojęcia | |
| Historia myśli ewolucyjnej |

