

Gatunek – w biologii podstawowa jednostka taksonomiczna, jedna z kategorii systematycznych oraz jednostka różnorodności biologicznej.
Ustalenie wyczerpującej definicji gatunku okazało się trudne. Próby takie są nadal podejmowane, ponieważ jest to pojęcie kluczowe dla praktyki badawczej naukowców i działań ruchu na rzecz ochrony przyrody. Karol Linneusz (a za nim wielu innych) przyjmował, że gatunki są czymś stałym, jednak obecnie wiadomo, że procesy ewolucyjne powodują ich przemiany i stopniowe przechodzenie jednych gatunków w inne. Często gatunek definiuje się jako największą grupę organizmów, spośród której dwa osobniki są w stanie rozmnożyć się (zazwyczaj płciowo), wydając płodne potomstwo. Jest to definicja ogólnie trafna, jednak błędna w wielu szczególnych przypadkach. Przykładowo nie znajduje ona zastosowania dla mieszańców, w kompleksach gatunków złożonych z setek podobnych mikrogatunków oraz gatunków pierścieniowych. W ich przypadku zaciera się granica między blisko spokrewnionymi gatunkami. Jest również niemożliwa do zastosowania w sytuacji rozmnażania bezpłciowego, czy w przypadku organizmów kopalnych, których rozrodu zbadać nie można. W paleontologii używa się więc koncepcji chronogatunku. Inne sposoby definiowania gatunku mogą odbywać się z pomocą badań kariotypów, sekwencji DNA, morfologii, zachowania czy nisz ekologicznych.
Każdy gatunek otrzymuje dwuczłonową (binominalną) nazwę. Pierwszy jej człon określa rodzaj, do którego należy, a drugi znany jest jako epitet gatunkowy bądź nazwa gatunkowa. W języku angielskim istnieje tu rozróżnienie na specific name i specific epithet (zwykle używane w nomenklaturze botanicznej i czasami zoologicznej). Np. Boa constrictor jest jednym z gatunków rodzaju Boa.
Od czasów Arystotelesa do XVIII wieku gatunek postrzegano jako coś stałego, o wyznaczonym miejscu w hierarchicznym wielkim łańcuchu bytu. W XIX w. biolodzy zrozumieli, że w odpowiednio długim czasie gatunki mogą ewoluować. W O powstawaniu gatunków Karola Darwina (1859) wyjaśniono, jak nowe gatunki powstają na drodze doboru naturalnego. Zrozumienie tego procesu pogłębiono w XX dzięki genetyce i ekologii populacyjnej. Zmienność genetyczna pojawia się wskutek mutacji oraz rekombinacji, natomiast same organizmy przemieszczają się, co prowadzi do izolacji geograficznej i dryfu genetycznego ze zmienną presją selekcyjną. Geny niekiedy są wymieniane między gatunkami dzięki poziomemu transferowi genów. Nowe gatunki mogą wyłonić się wskutek hybrydyzacji oraz poliploidii. Gatunki również wymierają z różnych powodów. Wirusy stanowią specjalny przypadek, jako że kształtuje je równowaga mutacyjno-selekcyjna, dzięki której występują w formie quasi-gatunków.
W praktyce wykorzystuje się różne definicje gatunku do mierzenia bioróżnorodności, jednak trafność tych pomiarów jest dyskusyjna.
Historia
Klasyczne formy
W swoim ujęciu biologii Arystoteles użył terminów γένος (génos), oznaczającego „rodzaj”, jak ptak czy ryba, oraz εἶδος (eidos) określającego „formę”, jak żuraw, orzeł, kruk czy wróbel (w obrębie „rodzaju” – ptaka). Choć słowa te przetłumaczono na łacinę jako „rodzaj” i „gatunek”, nie pokrywają się znaczeniowo z rodzajem i gatunkiem w taksonomii linneuszowskiej. Ptaki (Aves) są gromadą, żurawie (Gruidae) rodziną, a kruk (Corvus) rodzajem. Każdy z arystotelesowskich „rodzajów” miał pewne własności – tak jak ptaki mają pióra, dziób, skrzydła, twardą skorupę jaja i są stałocieplne. „Rodzaje” konstruowano poprzez szukanie istot mających każdą z tych cech, którą dodatkowo mogły w dowolnym układzie dziedziczyć młode. Arystoteles przekonany był, że wszystkie „rodzaje” i „formy” są wyraźnie zaznaczone i niezmienne. Pogląd ten panował aż do renesansu.
W różnego typu taksonomiach ludowych podstawowe wyróżniane taksony często odpowiadają rodzajowi, podczas gdy gatunki są wyróżniane epitetem. Przykłady takich ludowych to „rodzaj” róża i „gatunki” róża biała i róża czerwona czy „porzeczka” i „porzeczka czarna” oraz „czerwona”. Często jednak również w tym przypadku gatunki są określane nazwą jednoczłonową, np. w polskich taksonomiach ludowych gatunek borówka czarna rozpoznawany jest jako odrębny gatunek o nazwie, w zależności od regionu, „borówka” lub „jagoda”.
W nowożytnym przyrodoznawstwie pojawiło się podejście nominalistyczne, zgodnie z którym gatunek nie jest realnie istniejącym bytem, a jedynie określeniem pomagającym grupować podobne osobniki. W tym duchu Jean-Baptiste de Lamarck uważał, że osobniki, krzyżując się między sobą, dają potomstwo o coraz bardziej zatartych różnicach, aż do stopnia, gdy są tak podobne, że się ich nie odróżnia i zalicza do jednego gatunku.
Niezmienne gatunki

Gdy nowożytni obserwatorzy przyrody zaczęli rozwijać systemy klasyfikacji istot żywych, umieszczali każdy „rodzaj” zwierzęcia i rośliny w pewnym kontekście. Wiele z tych dawnych schematów klasyfikacji można dziś uznać za dziwaczne: były wśród nich wytyczne oparte na kolorach (wszystkie rośliny o żółtych kwiatach) czy zachowaniu (węże, skorpiony i pewne mrówki). John Ray, angielski przyrodnik, jako pierwszy podjął się sformułowania biologicznej definicji gatunku (1686):
Nie nasuwa mi się kryterium określania gatunku słuszniejsze od tego opartego na rozróżnianiu cech utrzymywanych [już] od nasiona poprzez rozród. Stąd nie ma znaczenia, jakie odchyły od normy pojawią się u osobników lub gatunków. Jeśli są zrodzone z nasiona jednej i tej samej rośliny, to stanowią przypadkową zmienność, nie na tyle [znaczącą], by uznać ją za gatunek ... Podobnie i zwierzęta różniące się między sobą zachowują swoje odrębne gatunki na stałe; jeden gatunek nigdy nie wyłoni się z nasienia innego i odwrotnie

W XVIII wieku szwedzki przyrodnik Karol Linneusz sklasyfikował organizmy ze względu na współdzielone przez nie cechy fizyczne, nie tylko różnice między nimi. Zakorzenił ideę taksonomicznej hierarchii w klasyfikacji organizmów, która oparta być miała na obserwowalnych cechach i docelowo powinna odzwierciedlać naturalne powiązania organizmów ze sobą. Ówcześnie powszechne było przekonanie, jakoby między gatunkami nie było naturalnych powiązań, niezależnie od ich podobieństwa. Stanowisko to wspierane było przez uczonych z tamtych lat i edukację religijną, wedle której kategorie, w których zawierają się organizmy żywe, dyktował Bóg. Ułożone były w arystotelesowskiej hierarchii, drabinie bytów. Choć nie wyrażono charakteru tych organizmów, stałego lub zmiennego, drabina zakłada możliwość wspięcia się. Inni autorzy, w tym Frédéric Cuvier, przekonani byli o sukcesywnym udoskonalaniu bytów przez Boga, przy czym każda próba zakończona była katastrofą. Za ostatnią z takich uznawał potop.
Możliwość zmiany
Gdy Linneusz zmierzył się z dowodami na hybrydyzację, zaakceptował możliwość zmiany gatunków i walkę o przetrwanie, ale nie swobodną ewolucję gatunków. Do XIX wieku przyrodnicy zrozumieli, że gatunki z czasem mogą się zmieniać, a czas istnienia Ziemi jest wystarczający do zajścia tych najważniejszych zmian. Jean-Baptiste Lamarck w swoim dziele Philosophie zoologique (1809) opisał transformizm. Sugerował, że gatunki mogą zmieniać się z czasem, co było drastycznie oderwane od idei arystotelesowskich.
W 1859 Karol Darwin i Alfred Russel Wallace dostarczyli przekonującego wyjaśnienia ewolucji i tworzenia się nowych gatunków. Darwin sądził, że ewoluowały populacje, nie jednostki, a działo się to na drodze doboru naturalnego; naturalnie występujących różnic między osobnikami. Wymagało to nowej definicji gatunku. Darwin wywnioskował, że gatunki są tym, czym się wydają: ideami, umożliwiającymi prowizoryczne nazwanie grupy osobników wchodzących ze sobą w interakcje (tłumaczenie w wersji Szymona Dicksteina):
Z powyższych uwag widać, że uważam słowo „gatunek” za nazwę nadawaną dowolnie, dla dogodności tylko, grupie jednostek mocno podobnych do siebie, że pojęcie to nie różni się istotnie od pojęcia „odmiana”, którym nazywa się formy mniej wyodrębnione i bardziej ulegające wahaniom. Podobnie i nazwy „odmiany” w porównaniu do różnic indywidualnych także używa się zupełnie dowolnie i tylko dla dogodności
Taksonomia i nazewnictwo

Nazwy zwyczajowe i naukowe
Pospolicie używane nazwy organizmów nierzadko mogą być mylące – przykładowo słowo „kot” oznaczać może kota domowego (Felis catus) lub dowolnego innego kotowatego (Felidae). Innym problemem jest zmienność nazw w zależności od regionu. W różnych częściach Ameryki puma płowa (Puma concolor) znana jest jako puma, kuguar, catamount, pantera, painter i lew górski. „Pantera” może jednak oznaczać również jaguara amerykańskiego (Panthera onca) z Ameryki Łacińskiej i lamparta plamistego (Panthera pardus) z Afryki i Azji. Nazwy naukowe powinny dla odróżnienia być unikatowe i uniwersalne. Składają się z nazwy rodzajowej, jak Puma, i epitetu gatunkowego, jak concolor. Pełna nazwa naukowa obejmuje również autora i rok pierwszego opisu, jak Locusta migratoria, Linneusz, 1758. Jeśli od pierwszego opisu zaszła zmiana nazwy naukowej nazwisko autora i rok obejmuje się okrągłym nawiasem. Dla przykładu, turkawka zwyczajna po raz pierwszy wspomniana została przez Linneusza jako Columba turtur, stąd podając obowiązującą nazwę Strepropelia turtur wspomniane dane należy zapisać: (Linneusz, 1758). Nazwiska autorów bywają zapisywane inaczej lub skracane, jak Karl von Linnaeus → Linnaeus → Linné. W przypadku niektórych grup organizmów lokalne środowiska naukowe publikują wykazy nazw gatunkowych w lokalnym języku utworzone na zasadach zbliżonych do nomenklatury naukowej.
Opisy gatunków

Każdy gatunek przy okazji opisu holotypu (czy też innego typu, w razie niedostępności holotypu) otrzymuje unikatową nazwę gatunkową. Nie zawsze da się jednoznacznie wskazać holotyp. Niektórzy XVIII-wieczni autorzy i większość XIX-wiecznych nie mieli w zwyczaju jednoznacznie wzmiankować jakiegoś okazu, który byłby holotypem. Wszystkie okazy, jakie autor miał przed oczami opisując nowy gatunek lub możliwe do rozpoznania w kolekcjach zwą się syntypami. Zdarza się również, że holotyp ginie. W razie przesłanek o odpowiedniej wadze (wynikającej z problemu natury biologicznej lub nomenklatorycznej) wskazuje się neotyp, przy czym należy zachować pewne kryteria, w tym pochodzenia (jak najbliższe miejscu pochodzenia holotypu). Przeważnie opis nowego gatunku zawiera informację o cechach pomocnych w identyfikacji przedstawiciela nowego gatunku, wyróżniających go na tle innych oraz spokrewnionych i podobnych organizmów, z którymi można takowego pomylić. Zapewnia tym samym źródło nazwy opublikowanej ważnie (w botanice, regulowana przez Międzynarodowy Kodeks Nomenklatury Botanicznej) lub nazwy przydatnej (nom utilise; regulowana przez Międzynarodowy Kodeks Nomenklatury Zoologicznej), jeśli zostanie zaakceptowany do publikacji.
Typy przechowuje się zwykle w trwałych przybytkach, nierzadko w kolekcjach służących do badań w znamiennych muzeach czy uniwersytetach. Ma to umożliwić dostęp do kolekcji pracującym nad daną grupą organizmów specjalistom oraz zapobiec niszczeniu zbiorów, co nierzadko miało miejsce w przypadku tych prywatnych, szczególnie skupiających owady i mięczaki. Autorzy pierwszego opisu gatunku zobowiązani są wybrać nazwę która, zgodnie z Kodeksem Nomenklatury Zoologicznej, jest właściwa, zwięzła, przyjemna dla ucha, pozostająca w pamięci i nieobraźliwa. Nazwy naukowe zawsze zaznacza się czcionką inną niż w otaczającym tekście, zazwyczaj kursywą, gdy większość tekstu pisana jest antykwą (lub odwrotnie). Według praktyki nazwy naukowe zapisuje się alfabetem łacińskim również tam, gdzie całość tekstu zapisana jest w innym, na przykład w cyrylicy czy alfabecie arabskim.
Skróty
W książkach i artykułach niekiedy celowo nie określa się gatunku z największą możliwą dokładnością, stąd używa się skrótów sp. (liczba pojedyncza) i spp. (liczba mnoga) dla określenia mnogości gatunków (species pluralis) w miejsce epitetu gatunkowego, np. Canis sp. Skrót ten nierzadko pojawia się w razie braku możliwości doprecyzowania gatunku, na przykład w paleontologii. Autorzy używają niekiedy skrótu spp. jako krótkiego komunikatu informującego o prawdziwości pewnego stwierdzenia względem wielu gatunków z rodzaju, ale nie wszystkich. Gdy owo twierdzenie prawdziwe jest względem przedstawicieli każdego z gatunków danego rodzaju, używa się samej nazwy rodzajowej. Nazwy rodzajów i gatunków powinny być zapisywane kursywą, w odróżnieniu od skrótu sp. i pokrewnych. W razie braku pewności w oznaczeniu danego osobnika wykorzystuje się skrót cf., umieszczany przed epitetem gatunkowym. Innymi abrewiacjami są nr. (w pobliżu) lub aff. (species affinis; gatunek podobny), którym autorzy posługują się w razie braku pewnej tożsamości danego osobnika, który jednak zdaje się być podobny do wymienionego później.
Kody identyfikacji
Wraz z pojawieniem się internetowych baz danych opracowano kody umożliwiające identyfikację już zdefiniowanych gatunków, w tym w:
- National Center for Biotechnology Information (NCBI), gdzie stosowany jest liczbowy taxid, czyli Taxonomy identifier, określony jako „stabilny i unikatowy identyfikator”; przykładowo, dla Homo sapiens jest to 9606.
- Kyoto Encyclopedia of Genes and Genomes (KEGG), w którym stosuje się trzy- lub czterocyfrowe kody określające ograniczoną gatunków; dla H. sapiens jest to hsa.
- UniProt zapewnia identyfikatory składające się z nie więcej, niż pięciu znaków alfanumerycznych, jak HUMAN dla H. sapiens.
- Integrated Taxonomic Information System (ITIS) zawiera unikatową liczbę dla każdego gatunku. Dla Homo sapiens LSID to urn:lsid:catalogueoflife.org:taxon:4da6736d-d35f-11e6-9d3f-bc764e092680:col20170225, a TSN (Taxonomic System Number) to 180092.
„Lumping” i „splitting”
Nazwanie konkretnego gatunku, w tym jego przynależności do określonego rodzaju (lub wyższego taksonu), jest równoznaczne z wysunięciem hipotezy związanej z pokrewieństwem ewolucyjnym i rozróżnialnością omawianej grupy organizmów. Wraz z napływem nowych informacji ta hipoteza może zostać potwierdzona albo odrzucona. Niekiedy, szczególnie w dawnych czasach bez łatwej komunikacji między systematykami, taksonomowie pracujący w odosobnieniu określają dwa odrębne organizmy odrębnymi gatunkami, później jednak okazują się reprezentować ten sam. Zazwyczaj w takiej sytuacji zachowywana jest stara nazwa, a młodsza w procesie synonimizacji staje się młodszym synonimem (junior synonym). Łączenie dwóch taksonów w jeden określane jest mianem lumping (ang. lump – łączyć, skupiać). Dzielenie istniejącego już taksonu na kilka, często nowych, nazywane jest splittingiem (ang. split – rozdzielać, rozszczepiać). Systematycy bywają przez współpracowników określani mianem lumpers lub splitters, zależnie od ich podejścia do wyszukiwania różnic bądź cech wspólnych między taksonami. Przykładowo w pewnej wydanej przez Uniwersytet Oksfordzki pozycji poświęconej pelikanowym, głuptakowym i pokrewnym Gregory Macalister Mathews określony został jako notorious splitter (notorious – notoryczny, znany [z czegoś]).
Szeroki i wąski zakres znaczeniowy
Kodeksy nomenklatury, takie jak Międzynarodowy Kodeks Nomenklatury Zoologicznej oraz Międzynarodowy Kodeks Nomenklatury Botanicznej, nie określają zasad ustalania granic gatunku. Badania mogą zmienić ich postrzeganie wraz z napływem nowych dowodów. Niekiedy zachodzi potrzeba wskazania granic, w jakich postrzegany jest dany gatunek, stąd używane są przeciwstawne określenia sensu stricto („w ścisłym znaczeniu”) oraz sensu lato („w szerokim znaczeniu”). Przykładowo fraza sensu lato może być użyta, gdy w tym ujęciu danego gatunku jest uwzględniony dodatkowy podgatunek. Inne skróty, w tym auct. (auctorum – „autor”) i non. („nie”) używane są do określenia autora, w myśl którego przedstawiono dany gatunek.
Koncepcja gatunku autorstwa Mayra

Większość współczesnych podręczników jako podstawę dalszego informowania o kwestii gatunku używa koncepcji gatunku biologicznego opracowanej przez Ernsta Mayra w 1942. Definicja brzmi:
grupa realnie lub potencjalnie krzyżujących się populacji, które rozrodczo odizolowane są od podobnych sobie grup
Podnoszone były argumenty, wedle których definicja ta stanowi naturalną konsekwencję wpływu rozmnażania płciowego na dynamikę selekcji naturalnej. Użycie przez Mayra przymiotnika potencjalny było przedmiotem sporów. Niektóre interpretacje wyłączają z tej definicji nietypowe albo przeprowadzone w sposób sztuczny krzyżowanie możliwe do uzyskania tylko w niewoli oraz dotyczące zwierząt mogących rozmnażać się, lecz nie czyniących tego w naturze. Mayr był jednym z największych specjalistów zajmujących się zagadnieniem gatunku. Swoją definicję modyfikował dalej, najbardziej w 1957 i 1969.
Problem koncepcji gatunku
Nie jest łatwe zdefiniowanie gatunku w sposób możliwy do zastosowania dla wszystkich organizmów. Trudności z tym związane są znane jako problem koncepcji gatunku. Dostrzeżono je już w 1859, kiedy Darwin zanotował (tłumaczenie w wersji Szymona Dicksteina):
Ani jedna definicja nie zadowoliła dotychczas wszystkich naturalistów, chociaż każdy z nich wie mniej więcej, co przez to słowo rozumieć. Zazwyczaj pojęcie to zawiera w sobie nieznany czynnik oddzielnego aktu stworzenia
Koncepcja Mayra – gdzie zawodzi?

Prostą, podręcznikową wręcz definicją Mayra można opisać gatunki organizmów wielokomórkowych. Nie znajduje jednak zastosowania dla następujących przypadków:
- Organizmy rozmnażające się bezpłciowo, jak jednokomórkowce (bakterie i inne prokarionty) oraz organizmy wielokomórkowe ze zdolnością partenogenezy lub apomiksji. Niekiedy używa się terminu quasi-gatunki do określania gwałtownie mutujących jednostek, jak wirusy.
- Brak naukowej wiedzy na temat możliwości krzyżowania się grup dwóch morfologicznie podobnych grup organizmów. Ma to miejsce w paleontologii, kiedy niemożliwe jest zbadanie zdolności rozrodczych danych organizmów.
- Znaczący przepływ genów między gatunkami za sprawą mieszańców.
- Gatunki pierścieniowe, w przypadku których przedstawiciele występujących blisko populacji są zdolni do rozrodu, ale przedstawiciele tych odległych nie.

Identyfikacja gatunków może nastręczać trudności wskutek rozbieżności wyników badań molekularnych i morfologicznych. Występują tu dwa problemy: zbieżna morfologia, lecz rozbieżne pochodzenie (konwergencja, gatunki bliźniacze) oraz rozbieżna morfologia, lecz wspólne pochodzenie (plastyczność fenotypowa czy różny wygląd na poszczególnych etapach rozwoju). Dodatkowo określenie gatunku utrudnia poziomy transfer genów (PTG). Większość definicji zakłada, że organizm potomny otrzymuje swoje geny od jednego lub dwóch rodziców o podobnych cechach, aczkolwiek w przypadku PTG tak się nie dzieje. Istnieją mocne dowody na zachodzenie PTG u niepodobnych do siebie grup eukariontów, w tym niektórych skorupiaków i szkarłupni.
Biolog ewolucyjny James Mallet stwierdził:
Nie ma łatwego sposobu na określenie, czy spokrewnione ze sobą geograficzne czy przejściowe formy należą do jednego, czy więcej gatunków. Luki między gatunkami ustalić można jedynie w skali lokalnej, o danym czasie. Jest się niejako zmuszonym przyznać rację Darwinowi: realność i niepodzielność gatunku będzie wysoce zredukowana w skali dużego terenu czy długiego czasu
Grupy mikrogatunków
Kolejne koncepcje gatunku podkopywane są przez istnienie mikrogatunków – grup organizmów, w tym wielu roślin, z niewielką zmiennością genetyczną, zazwyczaj tworzących kompleks gatunków. Przykładami takich grup są: mniszek pospolity (Taraxacum officinale; około 250 mikrogatunków tylko na Wyspach Brytyjskich) oraz jeżyna krzewiasta (Rubus fruticosus; około 300 mikrogatunków na Wyspach Brytyjskich). Co więcej, do stopnia skomplikowania systematyki wspomnianych organizmów przyczynia się występowanie hybrydyzacji, apomiksji i poliploidii. Kompleksy gatunków występują również u innych grup organizmów, jak motyle z rodzaju Heliconius, żaby Boana czy muchomor czerwony (Amanita muscaria).

Jeżyna, której owoce zjada człowiek, należy do jednego z kilkuset mikrogatunków w kompleksie gatunku Rubus fruticosus

Rodzaj motyli Heliconius zawiera wiele podobnych gatunków

Kompleks gatunków Hypsiboas calcaratus–fasciatus zawiera przynajmniej sześć gatunków rzekotek



Hybrydyzacja
Naturalnie występujące hybrydy stanowią wyzwanie dla koncepcji gatunku o zachowanej izolacji rozrodczej, jako że płodne mieszańce umożliwiają przepływ genów pomiędzy dwiema populacjami. Czarnowron (Corvus (corone) corone) i wrona siwa (Corvus (corone) cornix) bywają uznawane za odrębne gatunki, jednak swobodnie krzyżują się w miejscach nachodzenia się ich zasięgów. Występowanie mieszańców międzygatunkowych nie jest rzadkie w przyrodzie; przykładowo we florze Wielkiej Brytanii liczącej 2,5 tysiąca gatunków rozmnażających się płciowo zarejestrowano 780 takich mieszańców. Przypuszcza się, że od 30 do 70% gatunków w świecie roślin może mieć pochodzenie mieszańcowe.
- Krzyżowanie się wrony siwej i czarnowrona zapewnia przepływ genów między „gatunkami”

Hybryda z ciemnym brzuchem i ciemnoszarym karkiem

Hybryda z ciemnym brzuchem




Gatunki pierścieniowe
Gatunek pierścieniowy stanowi połączoną serię sąsiadujących ze sobą populacji. Zachowane są dwa warunki: przedstawiciele każdej z nich mogą krzyżować się z przedstawicielami przyległych populacji, niemniej istnieją przynajmniej dwie „krańcowe” populacje, zbyt sobie odległe, by ich reprezentanci mogli wydać płodne potomstwo. Te dwie niemożliwe do skrzyżowania, choć genetycznie połączone, populacje mogą współwystępować zamykając „pierścień”. Stanowią wyzwanie dla każdego konstruktu, który definicję gatunku opiera o izolację rozrodczą, jednak przykłady gatunków pierścieniowych są bardzo nieliczne. Należą do nich: kompleks mew Larus argentatus–L. fuscus (wokół bieguna północnego), grupa 19 populacji salamandry Ensatina eschscholtzii (Ameryka) oraz przedstawiciele Phylloscopus w Azji. Wiele spośród domniemanych gatunków pierścieniowych okazało się efektem błędnej klasyfikacji, co doprowadziło do spekulacji na temat istnienia takowych.

Na dwóch przeciwnych stronach pierścienia znajdują się mewy: srebrzysta (L. argentatus) oraz żółtonoga (L. fuscus)

Wójcik (Phylloscopus trochiloides)
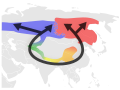
Prawdopodobna droga ewolucji ptaków z rodzaju Phylloscopus wokół Himalajów




Próby definiowania gatunku
Biolodzy i taksonomowie podejmowali liczne starania celem zdefiniowania gatunku, od podstaw morfologicznych po genetyczne. Linneusz i jemu współcześni dawni systematycy mogli jedynie opisywać to, co widzieli: później formalnie określono to mianem gatunku morfologicznego albo typologicznego. Mayr podkreślił rolę izolacji rozrodczej, acz bywa ona trudna lub niemożliwa do wykazania. Późniejsi biolodzy usiłowali udoskonalić definicję Mayra przez tworzenie nowych koncepcji gatunku – kohezyjną (spójności) i „rozpoznawania partnera”. Niektóre z tych pojęć są bardzo zbliżone, niekiedy nakładają się, stąd nie jest łatwo ustalić ich liczbę: biolog R. L. Mayden odnotował około 24 definicji gatunku, zaś filozof nauk John Wilkins już 26, a K. Lang 33 – opracowując w 1988 jedno z bardziej kompletnych zestawień. Wilkins pogrupował owe pojęcia gatunku w kilka grup: (1) „agamogatunki” dla organizmów rozmnażających się bezpłciowo (2) „gatunki biologiczne” dla organizmów izolowanych rozrodczo (3) „ekogatunki” oparte o nisze ekologiczne (4) gatunki ewolucyjne oparte o linię rozwojową (5) gatunki genetyczne wyodrębnione na podstawie puli genowej (6) gatunki morfologiczne oparte na fenotypach (7) gatunki taksonomiczne, czyli opisane tak, jak uznał to taksonom Natrafić można również na pogląd, wedle którego nie dysponujemy empirycznymi argumentami pomocnymi w odpowiedzi na pytanie o istotę gatunku, stąd też pozostaje ono pytaniem filozoficznym. Niektórzy autorzy próbują odwoływać się do uniwersalnej koncepcji gatunku, która jednak nigdy nie została opracowana i traktowana jest jako nieosiągalny ideał. W filozofii przyrody nominalizm zakłada, że gatunek nie jest rzeczywistym bytem, w odróżnieniu od osobników, a nazwa „gatunek” jedynie służy do określania osobników do siebie podobnych.
Niemniej jednak część taksonów występujących w przyrodzie uznawana bywa za „gatunki idealne”, stanowiąc wyraźnie odrębne i rozpoznawalne jednostki (nieciągłość fenotypu), izolowane genetycznie od innych gatunków (niezdolne do krzyżowania). Często takie gatunki występują w rodzinach roślin takich jak np. bobowate, selerowate, dzwonkowate.
Gatunek typologiczny i morfologiczny

Gatunek typologiczny stanowi grupa organizmów, których cechy podporządkować można jednemu ustalonemu zestawowi cech (typowi), stąd nawet osoby niepiśmienne mogą mieć podobne pojęcie takiego gatunku, jak współcześni systematycy. Tym rozumieniem gatunku posługiwali się już dawni myśliwi, rolnicy, wędkarze i lekarze, którzy musieli rozróżniać między organizmami szkodliwymi lub niebezpiecznymi a pożytecznymi. Zgrupowania odchyłów od normy lub zmienność fenotypowa w obrębie tego samego gatunku (na przykład długie i krótkie ogony) są tym, co w myśl koncepcji gatunku morfologicznego odróżnia organizmy z innych gatunków od siebie. Metoda ta stanowiła „klasyczną”, gdy przychodziło określać gatunek tak wcześnie, jak w czasach Linneusza. Różne fenotypy nie muszą jednak stanowić o przynależności do różnych gatunków, tak jak w przypadku czteroskrzydłych muszek Drosophila z dwuskrzydłych matek. Wyodrębniono więc gatunek morfologiczny. Systematyka topologiczna i ludowa może być jednak wysoce niedokładna. We Francji pod pojęciem „muchy” kryją się przedstawiciele około 4000 gatunków (stan z początku lat 90. XX wieku). Niekiedy jednak rdzenna ludność rozpoznaje zwierzęta z niesamowitą wręcz precyzją. Jared Diamond w 1965 opublikował badanie nad ludową taksonomią nowogwinejskiego plemienia Fore. Wówczas na obszarze znano 120 gatunków ptaków. Rdzennej ludności znane było 110, przy czym w 93 przypadkach ich nazwy gatunków dokładnie odpowiadały rozpoznaniu naukowców. W 4 przypadkach nazwy dla samców i samic były inne – dotyczyło to cudowronek, których pióra tubylcy wysoko sobie cenili. W 9 przypadkach nazwy nadano dwóm lub więcej gatunkom, które badacze uważali za wyjątkowo bliskie, lecz odróżnialne. Mieszkańcy zaprowadzeni poza swoje miejsca zamieszkania, poniżej 1500 m n.p.m., na 103 zaobserwowane gatunki nazwali swoimi nazwami 91, wszystkie były tymi najbliżej spokrewnionymi z pierwowzorem.
W latach 70. XX wieku Robert R. Sokal, Theodore J. Crovello i Peter Sneath zaproponowali nową odmianę istniejącej już koncepcji – gatunek fenetyczny. Miałby określać grupę osobników z podobnymi fenotypami względem siebie, ale odmiennymi fenotypami w porównaniu do innych grup gatunków. Różnica między gatunkiem morfologicznym a fenetycznym polega na zawarciu w definicji tego drugiego pewnych wartości numerycznych uzyskanych po zbadaniu grup jednostek. Uzyskuje się je na drodze porównania wielu zmiennych uzyskanych z odpowiednio dużej grupy danych fenetycznych. W systematyce numerycznej, zwanej też fenetyczną, rezultaty przedstawia się na dendrogramach. Nie mają one „korzeni”, jako że nie odzwierciedlają procesów zachodzących w wymiarze czasu. Odległości symbolizują pokrewieństwo gatunków ustalone przez komputerową analizę cech wspólnych – od 0 (brak badanych cech wspólnych) po 1000 (całkowita zgodność). Można przyjąć, że wartości numeryczne odpowiadać będą rangom – na przykład założyć, że próg 800 symbolizował będzie rodzaj. Najprostszy dendrogram będzie obrazował rozkład dwóch zmiennych, można również dodając trzecią przenieść dendrogram na poziom trójwymiarowy i tak dalej. Teoretycznie fenetyka pod uwagę bierze maksimum dostępnych cech, nie zważając na ich naturę.
Kryterium spójności i rozpoznawania partnera
Idea gatunku opartego o rozpoznawanie partnerów zakłada, że grupa rozmnażających się płciowo organizmów rozpoznaje inne jako potencjalnych partnerów. Rozszerzając to kryterium o możliwość izolacji po sezonie rozrodczym, dochodzi się do kryterium spójności (kohezji): to największa populacja, w której osobniki mają potencjał grupowania się według fenotypów przez własny „wewnętrzny napęd”. Nie ma znaczenia, czy mogą z powodzeniem krzyżować się – nadal spełniają kryterium spójności, jeśli hybrydyzacja nie wystarczy do wymieszania ich genów z genami innej populacji. Rozwinięciem tego ujęcia jest biosemiotyczna koncepcja gatunku.
Podobieństwo i metkowanie genetyczne

W mikrobiologii geny mogą dość swobodnie przemieszczać się nawet między odlegle spokrewnionymi bakteriami, sięgając być może do całej domeny prokariontów. Z tego powodu mikrobiolodzy założyli, że bakterie lub archeony z sekwencjami genu kodującego podjednostkę rybosomu 16S rRNA są bardziej podobnymi, niż w 97% do siebie nawzajem wymagają przeprowadzenia hybrydyzacji DNA-DNA dla dalszego ustalenia przynależności. W 2006 zmieniono tę wartość na 98,7%. Sekwencje genu wspomnianej podjednostki mogą, prócz ustalania pokrewieństwa, służyć do identyfikacji samego gatunku pobranej próbki bakterii.
Hybrydyzacja DNA-DNA jest metodą przestarzałą i prowadziła do błędnych wniosków, jak w przypadku wydrzyka tęposternego (Stercorarius pomarinus) i wydrzyka wielkiego (Stercorarius skua). Nowoczesne podejścia uwzględniają porównywanie podobieństwa sekwencji z pomocą komputerowych obliczeń.
Metkowanie genetyczne (ang. DNA barcoding) przedstawiono jako metodę rozróżniania gatunków możliwą do wykorzystania nawet dla osób spoza kręgu specjalistów. Ta biologiczna metka, dosłownie „genetyczny kod kreskowy”, jest regionem mtDNA, na którym znajduje się gen kodujący oksydazę cytochromu c. Baza Barcode of Life Data Systems (BOLD) zawiera metki genetyczne dla ponad 190 tysięcy gatunków. Niektórzy naukowcy, jak Rob DeSalle, zwracają uwagę na konieczność pogodzenia klasycznej taksonomii z metkowaniem (które uznają za błędne określenie) ze względu na inaczej postrzegane granice gatunku. Introgresja wskutek działania endosymbiontów i innych wektorów może uczynić metkowanie nieskutecznym w identyfikacji gatunku.
Gatunek filogenetyczny (ewolucyjny)

Gatunek filogenetyczny lub kladystyczny reprezentuje ewolucyjnie odrębną linię rozwojową, która zachowała swoją dziedziczną integralność w czasie i przestrzeni. Stanowi najmniejszą grupę populacji, które rozróżnić można pośród innych przez zestaw unikalnych morfologicznych i genetycznych cech. Do ustalania genetycznego podobieństwa w mtDNA i nDNA służą markery genetyczne. Dla przykładu w przypadku grzybów właśnie badanie nukleotydów i rozpatrywanie gatunku w ujęciu kladystycznym przynosi najlepsze efekty w rozpoznawaniu gatunków akurat w tej grupie. Poszczególne wersje koncepcji gatunku filogenetycznego mogą skupiać się na monofiletyczności lub diagnozowalności.
W odróżnieniu od gatunku biologicznego, idea gatunku filogenetycznego nie jest wsparta o izolację rozrodczą, stąd funkcjonuje niezależnie od procesów nieodłącznych dla innych koncepcji. Znajduje zastosowanie dla organizmów bezpłciowych i może uwzględnić niedawne zmiany, czego pojęcia gatunku oparte o morfologię zrobić nie mogą. Nie w każdym przypadku zamysł ten będzie użyteczny, stąd niekiedy do uzyskania wyniku należy zbadać więcej niż jeden polimorficzny locus. Wdrożenie pojęcia gatunku filogenetycznego może doprowadzić do rozbicia istniejących gatunków na wiele innych, jak to miało miejsce w rodzinie wołowatych (Bovidae).
Gatunek ewolucyjny taki, jakim go opisał George Gaylord Simpson w 1951, stanowi byt utworzony z organizmów zachowujących swą odrębność względem innych im podobnych w czasie i przestrzeni, które przejawiają odmienne ewolucyjne przeznaczenie i tendencje historyczne. Od biologicznej koncepcji gatunku odróżnia go uwzględnienie trwałości na przestrzeni czasu. Wiley i Mayden twierdzili, że idea gatunku ewolucyjnego jest „tożsama” z tą opracowaną przez Williego Henniga, ukazującą gatunki jako linie rozwojowe. Hennig przyjmował cezurę czasową – istnienie gatunku musiało być rozpoczęte i zakończone przez specjację. Wspomniane koncepcje gatunków Wiley i Mayden uznali za „kilka wersji” tej filogenetycznej. Do tego ich zdaniem w naukach przyrodniczych niewłaściwe jest używanie takiej definicji gatunku, którą można zastosować i do wyższych taksonów, jako że w ich intencji leży dokładne poznanie liczby gatunków. Według wspomnianych biologów omawiane ujęcie gatunku obejmuje organizmy rozmnażające się i płciowo, i bezpłciowo. W myśl koncepcji ewolucyjnej taksony wyższe od gatunku rodzą się w momencie powstania pierwszego gatunku reprezentującego chociażby nową rodzinę.
Ekologiczna koncepcja gatunku
Gatunek ekologiczny stanowi grupę organizmów przystosowanych do zajmowania określonej niszy. Według tej koncepcji populacje tworzą fenetyczne zgrupowania, które postrzegane są jako gatunki, gdyż procesy ekologiczne i ewolucyjne kontrolujące rozdział dóbr sprawiają, że owe zgrupowania powstają.
Gatunek genetyczny
Gatunek genetyczny w myśl Roberta Bakera i Roberta Bradleya oznacza zbiór genetycznie izolowanych populacji osobników wzajemnie się krzyżujących. Jest to podobne do koncepcji gatunku biologicznego Mayra, jednak podkreśla znaczenie izolacji genetycznej, nie rozrodczej. W XXI wieku pojawiła się możliwość porównywania sekwencji DNA, ale wcześniej znano inne metody polegające na zestawianiu ze sobą kariotypów czy allozymów.
Jednostki ważne z punktu widzenia procesów ewolucyjnych
Jednostki ważne z punktu widzenia procesów ewolucyjnych (evolutionarily significant unit, ESU) lub „wildlife species” (dosł. gatunki dzikiej przyrody) są populacjami organizmów uznawanych za znaczące z punktu widzenia ochrony przyrody.
Chronogatunek
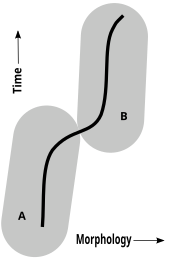
W paleontologii, gdzie dostępny jest jedynie morfologiczny materiał porównawczy ze szczątków fosylnych, stosuje się pojęcie chronogatunku. W anagenezie (formie ewolucji, w której nie dochodzi do rozgałęzienia i powstania nowej linii rozwojowej) paleontolodzy doszukują się sekwencji gatunków, gdzie każdy wywiedziony został z innego, już wymarłego, na drodze powolnych i mniej lub bardziej równomiernych zmian. Wyzwaniem dla paleontologów jest określenie ile zmian morfologicznych w danej jednostce czasu musi zajść, by można uznać danego osobnika za reprezentującego inny gatunek, niż jego przodek. Granice między chronogatunkami są rozmyte, co objawia się płynnymi przejściami w zapisie stratygraficznym.
Quasi-gatunki wirusowe
Wirusy mają niezmiernie liczne populacje i nie można jednoznacznie określić ich mianem „żywych”, jako że składają się niemal wyłącznie z nici DNA lub RNA w białkowej otoczce i gwałtownie mutują. Te cechy czynią konwencjonalne definicje gatunków w większości bezużytecznymi wobec wirusów. Quasi-gatunki wirusowe stanowią grupy genotypów powiązanych ze sobą przez podobne mutacje, konkurujące ze sobą w wysoce mutagennym środowisku i stąd rządzone przez równowagę mutacyjno-selekcyjną. Przewiduje się, że quasi-gatunek wirusowy w płaskim i nisko położonym krajobrazie dostosowania (ang. fitness landscape) wyprze quasi-gatunek położony wyżej, ale w węższej lokalizacji – na „szczycie” w krajobrazie dostosowania, wśród niedostosowanych mutantów. Zwie się to w języku angielskim survival of the flattest. Nie ma przesłanek, wedle których definicja gatunku wirusów byłaby podobna do tradycyjnych, biologicznych koncepcji gatunku.
Zmiany
Gatunki podlegają zmianom, zarówno poprzez ewoluowaniu w kolejne, wymianę genów, łączenie się z innymi, jak i wymieranie.
Specjacja
Proces ewolucyjny, w którym biologiczna populacja ewoluuje, by stać się znaczącą lub rozrodczo izolowaną zwie się specjacją. Karol Darwin jako pierwszy opisał rolę doboru naturalnego w specjacji w swoim dziele O powstawaniu gatunków (1859). Specjacja oparta jest o miarę izolacji rozrodczej, zredukowanego przepływu genów. Najłatwiej zmiana ta zachodzi poprzez specjację allopatryczną, w której czasie populacje są oddzielone barierą geograficzną i mogą stopniowo rozdzielać się wraz z akumulacją kolejnych mutacji. Izolację rozrodczą zakłócić może hybrydyzacja, dzieje się tak jednak tylko do momentu zaistnienia niekompatybilności alleli tego samego genu, co opisuje model Dobzhansky’ego-Mullera. Odmienny mechanizm, specjacja filetyczna, dotyczy dwóch linii, które stopniowo zlewają się tworząc nową formę, bez wzrostu liczby gatunków.
Międzygatunkowa wymiana genów
Poziomy transfer genów pomiędzy organizmami odmiennych gatunków, zarówno przez krzyżowanie, skok antygenowy, jak i reasortację genową stanowi niekiedy znaczące źródło zmienności genetycznej. Wirusy są w stanie wymieniać geny pomiędzy gatunkami. Bakterie mogą wymieniać plazmidy, w tym z bakteriami gatunków mocno od siebie odległych na drzewach filogenetycznych, co utrudnia ustalanie ich pokrewieństwa i osłabia koncepcje gatunku bakteryjnego.
Louis-Marie Bobay i Howard Ochman po zbadaniu genomów wielu bakterii zasugerowali, że mogą one zostać połączone w „grupy regularnie wymieniające geny”, na podobnej zasadzie, na jakiej zwierzęta i rośliny są łączone w grupy organizmów rozrodczo izolowanych. Bakterie mogą więc formować, analogicznie do biologicznego ujęcia gatunku Mayra, bezpłciowo rozmnażające się populacje wymieniające geny poprzez rekombinację.
Wymieranie
Wymieranie następuje w momencie śmierci ostatniego osobnika danego gatunku, który mógł jednak już wcześniej być wymarłym na poziomie funkcjonalnym. Szacuje się, że wymarło blisko 99% z 5 mld gatunków, które kiedykolwiek zasiedlały Ziemię. Niektóre ginęły podczas masowych wymierań (np. pod koniec permu, triasu i kredy), powodowanych przez różne przyczyny, jak aktywność wulkaniczna, zmiany klimatu, zmiany w składzie chemicznym oceanów i atmosfery. Wyraźnie odbiły się one na ekologii, atmosferze, powierzchni ziemi i wodach. Inną formą wymarcia jest „przyswojenie” jednego gatunku przez drugi przez hybrydyzację; w języku angielskim tak powstały gatunek zwie się compilospecies.
Praktyczne zastosowania koncepcji gatunku
Biolodzy i działacze na rzecz ochrony środowiska muszą identyfikować i kategoryzować organizmy, by umożliwić sobie pracę. Trudność, z jaką przychodzi wiarygodne przypisywanie organizmów do właściwych taksonów, stanowi zagrożenie dla wiarygodności ich prac, przykładowo podczas ustalania liczebności przedstawicieli danego gatunku w ekosystemie. Paul Michael-Agapow i współpracownicy ustalili, że użycie filogenetycznej koncepcji gatunku przyniosło im rozpoznanie o 48% więcej gatunków i odpowiednio mniejszych populacji, niż przy zastosowaniu innych ujęć. Według nich zjawisko to, nazwane „taksonomiczną inflacją”, może poskutkować zafałszowaniem liczby faktycznie zagrożonych gatunków i przynieść trudności polityczne i praktyczne. Biolog ewolucyjny Jody Hey zaobserwował wrodzony konflikt między potrzebą zrozumienia procesu specjacji a potrzebą identyfikacji i kategoryzacji. Prawa dotyczące ochrony przyrody w wielu państwach gwarantują zapobieganie wymieraniu gatunków. Strefy hybrydyzacji dwóch gatunków, objętego i nieobjętego ochroną, mogą prowadzić do konfliktów między prawodawcami, właścicielami danych terenów i działaczami na rzecz ochrony przyrody. Przykładem takiego nachodzenia się zasięgów prowadzącego do problemów z prawodawstwem jest krzyżowanie się chronionego puszczyka stokowego (Strix occidentalis caurina) z nieobjętym specjalną ochroną puszczykiem kreskowanym (Strix varia).
Botanik Brent D. Mishler twierdzi, podobnie jak Hey, że problem gatunku wykreowany został przez odmienne sposoby, na jakie ludzie chcą wykorzystywać definicje gatunku. Zaleca porzucenie tradycyjnych rang i używanie pojęcia grup monofiletycznych o różnej inkluzywności. Gatunek jest „po prostu najmniej inkluzywnym taksonem” dowolnego typu. Argumentuje, że praca nad ekologią, ewolucją i ochroną środowiska z takim ujęciem gatunku będzie lepiej funkcjonować. Jako że w tym ujęciu gatunków porównywać nie można, liczenie ich nie stanowi miarodajnego wskaźnika bioróżnorodności. Mishler proponuje więc, w myśl innych autorów jak R. I. Vane-Wright i współpracownicy, mierzenie bioróżnorodności poprzez liczbę rozgałęzień i prawdopodobnie również ich długość, oddzielającą koniuszki na drzewie [życia]. Mierzenie bioróżnorodności ma dodatkowe szczególne znaczenie – budzi zainteresowanie społeczeństwa, do którego świadomości za pośrednictwem mediów przenikają różne koncepcje gatunku.
Szacowanie liczby gatunków
Globalna bioróżnorodność zastanawiała biologów od wieków. John Ray w XVII wieku napisał, że w związku z odkryciami licznych ciem i motyli na terenie Wielkiej Brytanii liczba gatunków owadów w całym kraju może wynosić nawet 2000, a na całym świecie 20 tys. Karol Linneusz w swoim dziele Systema Naturae opisał blisko 4400 gatunków i przekonany był, że to liczba bliska globalnej bioróżnorodności. Przez kolejne dwa stulecia znana liczba gatunków znacznie powiększyła się, przynosząc nowe wyzwania dla biologów. Do lat 80. XX wieku uznawano, że globalna liczba gatunków jest 2–3 razy większa od liczby opisanych (np. 2–3 mln). Dyskusję nad szacowaniem liczby gatunków wznowiono, gdy w 1982 Terry Erwin ogłosił, że wedle jego szacunków gatunków na świecie jest blisko 30 mln. Wyliczenia opierały się na badaniu rozmieszczenia chrząszczy. Kolejne szacunki i proporcje tworzących tę bioróżnorodność grup zmieniały się:
- Wilson, 1992: 0,4% bakterie, 4,1% protisty, 17,6% rośliny, 4,9% grzyby, 73,1% zwierzęta,
- Mora et al., 2011 (nie uwzględnia w wyliczeniach prokariota w sposób wystarczający): ~8,7 mln gatunków, w tym ~2,2 mln morskich; w tym: 0,1% bakterie, 1% protisty, 2,8% rośliny, 5,6% grzyby, 90,5% zwierzęta,
- Larsen et al., 2017: ~2 miliardy gatunków, w tym 78% bakterie, 7,3% protisty, 0,02% rośliny, 7,4% grzyby, 7,3% zwierzęta. Badacze wzięli pod uwagę nowe informacje dotyczące owadów, których gatunków może być 40 milionów. Oszacowali również, że każdy gatunek zwierząt może być nosicielem dla przynajmniej 10 gatunków bakterii niespotykanych nigdzie indziej i stąd uzyskali owe 2 miliardy.
Bibliografia
- Loic Matile, Pascal Tassy, Daniel Goujet: Wstęp do systematyki zoologicznej: koncepcje, zasady, metody. Warszawa: PWN, 1993, s. 63.
- Claridge, H.A. Dawah, M.R. Wilson: Species. The units of biodiversity. Chapman & Hall, 1997. (ang.).Sprawdź autora:1.
- Quentin D. Wheeler, Rudolf Meier: Species concepts and phylogenetic theory: a debate. Columbia University Press, 2000. ISBN 978-0-231-10143-1. (ang.).
- Frank E. Zachos: Species concepts in biology. Historical Development, Theoretical Foundations and Practical Relevance. Springer International Publishing, 2016. (ang.).
Linki zewnętrzne
- Barcoding of species
- Catalogue of Life
- Definicja gatunku w Stanford Encyclopedia of Philosophy. plato.stanford.edu-online-documents-archive.info. [zarchiwizowane z tego adresu (2014-04-07)].
- VisualTaxa
- Wikispecies – wolny projekt Wikimedia Foundation