
| Rodentia | |||||
| Bowdich, 1821 | |||||
| Okres istnienia: 56–0 mln lat temu | |||||
 Na górze: kapibara wielka (jeżozwierzowce), postrzałka kafryjska (wiewiórolotkowe), pięknosuseł złocisty (wiewiórkokształtne), na dole: bóbr kanadyjski (bobrokształtne), mysz domowa (myszokształtne) | |||||
| Systematyka | |||||
| Domena | |||||
|---|---|---|---|---|---|
| Królestwo | |||||
| Typ | |||||
| Podtyp | |||||
| Nadgromada | |||||
| Gromada | |||||
| Podgromada | |||||
| Infragromada | |||||
| Rząd |
gryzonie |
||||
| |||||
| Podrzędy | |||||
| |||||
Gryzonie (Rodentia, od łacińskiego rodere, obgryzać) – rząd ssaków, charakteryzujący się pojedynczą parą ciągle wzrastających siekaczy zarówno w szczęce, jak i w żuchwie. Zalicza się do nich około 40% gatunków ssaków. Występują w dużej liczbie na wszystkich kontynentach z wyjątkiem Antarktyki. Stanowią najbardziej zróżnicowany rząd ssaków. Spotyka się je w różnorodnych siedliskach lądowych, w tym antropogenicznych. Obejmują gatunki nadrzewne, drążące w glebie i częściowo wodne. Zaliczają się do nich tak dobrze znane zwierzęta, jak myszy, szczury, wiewiórki, piesek preriowy, jeżozwierzowate, bobry, kawia domowa i chomiki. Niegdyś włączano do nich także i dziś często mylone z nimi zające, króliki i szczekuszkowate. Obecnie umieszcza się je w osobnym rzędzie zajęczaków.
Większość gryzoni ma dobrze zbudowane ciało, krótkie kończyny i długi ogon, jednak istnieją wyjątki. Zwierzęta te wykorzystują swoje ostre siekacze do gryzienia pokarmu, kopania jam i obrony. Większość żywi się nasionami lub innym pokarmem roślinnym, niektóre mają bardziej zróżnicowaną dietę. Wykazują tendencję do zachowań społecznych. Wiele gatunków żyje w społecznościach, w których członkowie w złożony sposób komunikują się ze sobą. Może wśród nich występować monogamia, poligamia aż do promiskuityzmu. U wielu (np. chomików) na świat przychodzi miot niedorozwiniętych, wymagających opieki w gnieździe młodych, podczas gdy inne (jak kawia domowa) rodzą młode od początku dobrze rozwinięte.
Skamieniałości gryzoni datuje się na czas od paleocenu, najstarsze pochodzą z Laurazji. W eocenie zróżnicowały się znacznie i rozprzestrzeniły na różne kontynenty, czasami nawet udało im się przebyć oceany. Z Afryki przedostały się do Ameryki Południowej i na Madagaskar. Jako jedyne lądowe łożyskowce dostały się na Australię i skolonizowały ją.
Gryzonie są wykorzystywane jako źródło mięsa, ich skóry są wykorzystywane do wytwarzania ubrań, służą też jako zwierzęta domowe lub laboratoryjne. Pewne gatunki, jak szczur śniady, szczur wędrowny czy mysz domowa, stały się poważnymi szkodnikami. Zjadają i niszczą żywność magazynowaną przez człowieka, roznoszą choroby. Przypadkowe wprowadzenie gryzoni do środowiska często czyni z nich gatunki inwazyjne, często zagrażające przetrwaniu gatunków rodzimych, jak zasiedlające wyspy ptaki, wcześniej izolowane morzem od lądowych drapieżników.
Budowa
Masa ciała największego współczesnego przedstawiciela gryzoni, kapibary wielkiej, może osiągać nawet 66 kg. Jednak masa ciała większości gryzoni nie przekracza 100 g. Najmniejszym gryzoniem jest trąbkouszek beludżystański (Salpingotulus michaelis), którego tułów wraz z głową osiąga 3,6–4,7 cm długości (ogon 7,2–9,4 cm), a tylna kończyna ma długość 1,8–1,9 cm, zaś masa dorosłej samicy osiąga tylko 3,75 g. Morfologia poszczególnych gryzoni jest zróżnicowana, ale typowo są zwierzętami naziemnymi, mają krępe ciało o długim ogonie i krótkich kończynach. Gryzonie ziemnowodne rozwinęły spłaszczenie – grzbietobrzuszne, spotykane u bobrów, bądź boczne, obserwowane u piżmaków. U form żyjących pod ziemią ogon uległ skróceniu. Ich kształt ciała określa się jako wałeczkowaty, a małżowiny uszne i oczy są w zaniku.

Gryzonie generalnie dysponują dobrze rozwiniętym zmysłem węchu, słuchu i wzroku. Gatunki nocne często mają duże oczy, niektóre rejestrują długości fal charakterystyczne dla ultrafioletu. Wiele z nich ma duże, czułe wibryssy. Niekiedy występują worki policzkowe, które mogą być pokryte futrem. Worki te mogą być wywracane na zewnątrz i czyszczone. U wielu gatunków język nie sięga za siekacze. Gryzonie mają wydajny układ pokarmowy. Pobierają prawie 80% dostarczanej energii. Zjadając celulozę, zmiękczają pokarm w żołądku, po czym przechodzi on do kątnicy, gdzie bakterie rozkładają go do prostszych węglowodanów. Gryzonie praktykują koprofagię – zjadają bryłki swojego kału, po czym zawarte w nich składniki odżywcze są wchłaniane przez jelito. Przeto gryzonie często tworzą twarde i suche grudki kału.
Gryzonie wykazują ogromne zróżnicowanie cech, nawet wśród blisko ze sobą spokrewnionych gatunków. Cechy kilku przedstawicieli tej grupy wymienia poniższa tabela.
| Gatunek | Maksymalna długość życia w latach († w niewoli) | Masa dorosłego osobnika w gramach | Ciąża w dniach | Liczba miotów w roku | Liczność miotu – średnia (i zakres) |
|---|---|---|---|---|---|
| mysz domowa (Mus musculus) | 4,0† | 20 | 19 | 5,4 | 5,5 (3-12) |
| golec piaskowy (Heterocephalus glaber) | 31,0 | 35 | 70 | 3,5 | 11,3 |
| szczur śniady (Rattus rattus) | 4,† | 200 | 21 | 4,3 | 7,3 (6-12) |
| szczur wędrowny (Rattus norvegicus) | 3,8† | 300 | 21 | 3,7 | 9,9 (2-14) |
| wiewiórka pospolita (Sciurus vulgaris) | 14,8† | 600 | 38 | 2,0 | 5,0 (1-10) |
| szynszyla mała (Chinchilla lanigera) | 17,2† | 642 | 111 | 2,0 | 2,0 (1-6) |
| kawia domowa (Cavia porcellus) | 12,0 | 728 | 68 | 5,0 | 3,8 (1-8) |
| nutria amerykańska (Myocastor coypus) | 8,5† | 7.850 | 131 | 2,4 | 5,8 (3-12) |
| kapibara wielka (Hydrochoerus hydrochaeris) | 15,1† | 55.000 | 150 | 1,3 | 4,0 (2-8) |

Czaszka
Czaszka gryzoni opisywana jest jako niska, wydłużona, spłaszczona z góry, o niewielkiej puszce mózgowej, w przeciwieństwie do okazałej jamy nosowej.
Oczodół gryzoni jest otwarty. Występuje duża, szczególnie u form zamieszkujących pustynie, puszka bębenkowa.
Kość nosowa jest wydłużona. Nozdrza przednie kierują się do przodu bądź do dołu.
Przez kość szczękową przebiega duży kanał podoczodołowy. Może w nim znajdować się fragment żwacza.
Kość zębowa prawa i lewa, dobrze rozwinięte, nie zrastają się w jedną kość, łączy je więzadło, co zwiększa elastyczność żuchwy, zdolnej do ruchów w kierunku przednio-tylnym oraz na boki. Występuje wyrostek kątowy, którego budowa stanowi podstawę podziału gryzoni na 2 grupy, zaproponowane przez Tulberga i odgrywające w przeszłości znaczenie w systematyce gryzoni:
- Gryzonie sciurognatyczne mają wyrostek kątowy leżący w przedłużeniu brzegu dolnego kości zębowej. Może on wystawać do wewnątrz.
- Gryzonie hystrikognatyczne mają wyrostek położony bardziej na zewnątrz.
Szwy czaszkowe istnieją w rozwoju długo.
Uzębienie
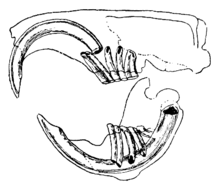
Cechę wyróżniającą gryzonie stanowi obecność ciągle rosnących, przypominających w kształcie brzytwę siekaczy. Przednią ich powierzchnię pokrywa gruba warstwa szkliwa, tylną zaś cienka. Ich zębodoły leżą w kościach międzyszczękowych. Ponieważ siekacze nie przestają rosnąć, zwierzę musi stale je ścierać, by nie sięgnęły ani nie przebiły kości czaszki. Gdy siekacze zwierają się ze sobą, bardziej miękka zębina tylnej powierzchni zębów ściera się, pozostawiając ostrą krawędź szkliwa jak brzeszczot czy dłuto. Większość gatunków posiada do 22 zębów, brakuje im jednak kłów i przednich przedtrzonowców. U większości gatunków siekacze i zęby policzkowe dzieli przerwa zwana diastemą. Pozwala im to ssać coś w policzku lub wargach, chroniąc pysk i gardło przed wiórami drewna bądź innym niejadalnym materiałem, a następnie pozbyć się odpadków bokiem przez usta. Szynszyle i kawie domowe żywią się pokarmem o dużej zawartości włókien. Ich trzonowce nie mają korzeni i rosną ciągle jak kły.
U wielu gatunków trzonowce są względnie duże, o skomplikowanej budowie, pokryte wieloma guzkami bądź grzbietami, jednakże u niektórych, jak Pseudohydromys, są mniejsze i prościej zbudowane. Są dobrze przystosowane do miażdżenia pokarmu na drobne porcje.
Aparat gębowy
Szczęka i żuchwa są silne umięśnione. Żuchwa podczas gryzienia wypychana jest ku przodowi, podczas żucia zaś pociągana ku tyłowi. Różne grupy gryzoni różnią się ułożeniem mięśni szczęki i żuchwy, jak też budową związanych z nimi struktur czaszki, tak między sobą, jak też i od innych ssaków. Wiewiórkokształtne, jak wiewiórka szara, posiadają duży, gruby mięsień żwacz, pozwalający im wydajnie gryźć siekaczami. Myszokształtne, jak szczur wędrowny, mają powiększony mięsień skroniowy, co czyni je zdolnymi do silnego żucia z udziałem trzonowców. Jeżozwierzowce, jak kawia domowa, dysponują większym powierzchownym mięśniem żwaczem, a mniejszym głębokim, niż szczury czy wiewiórki, wobec czego prawdopodobnie mniej wydajnie gryzą swymi siekaczami, ale ich powiększony mięsień skrzydłowy przyśrodkowy może umożliwiać im poruszanie żuchwą bardziej na boki przy żuciu. Wargi zbudowane są tak, że przy zamkniętych ustach siekacze wystają na zewnątrz, co umożliwia choćby rycie w ziemi. U niektórych gatunków występują torby policzkowe, w których zwierzę przetrzymuje pokarm. Mogą być one owłosione. W usunięciu z nich pożywienia gryzoń pomaga sobie kończynami przednimi.
Szkielet osiowy pozaczaszkowy
Gryzonie skaczące charakteryzują się skróceniem szyi. Ich kręgi zlewają się w tym odcinku kręgosłupa.
Kończyny i poruszanie się
U form skaczących śródstopie może zrastać się w pojedynczą kość.
Przednia para łap kończy się zazwyczaj pięcioma palcami, do których zalicza się przeciwstawny kciuk. Staw łokciowy zapewnia przednim kończynom znaczną giętkość. Tylne kończyny mają od trzech do pięciu palców, najczęściej jednak pięć. Zmniejszenie liczby palców może wynikać z redukcji palców bocznych, co obserwuje się zwłaszcza u gryzoni skaczących. U większości gatunków występuje stopochodność – chodzą na całej powierzchni dłoni czy stopy wraz z palcami, zakończonymi pazurowatymi paznokciami. Paznokcie gryzoni kopiących w ziemi są duże i mocne. Natomiast gryzonie nadrzewne mają paznokcie krótsze i ostrzejsze. U form prowadzących ziemnowodny tryb życia pomiędzy palcami może występować błona pławna.
Gryzonie wykorzystują szeroką gamę środków przemieszczania się. Zalicza się do nich chód czworonożny, bieg, drążenie w ziemi, wspinaczka, skoki dwunożne (szczuroskoczki, skakuszki), pływanie, a nawet szybowanie. Wiewiórolotkowate i polatuchy (nie są one ze sobą blisko spokrewnione) potrafią przemieścić się lotem szybowcowym z drzewa na drzewo, wykorzystując rozpościerające się pomiędzy przednimi a tylnymi kończynami błony. Aguti (Dasyprocta) są szybkonogimi palcochodami, a ich ruchy wykazują podobieństwo do sposobu poruszania się antylop. Ich palce kończą struktury przypominające kopyta. Gryzonie zasiedlające otwarte tereny z pustyniami włącznie przystosowały się do szybkiego poruszania się skokami, co ma związek ze znacznie lepszym rozwojem kończyn tylnych, niż przednich. Ponadto ich boczne palce stóp uległy redukcji.
Ogon
Większość gryzoni ma ogon, przybierający różne kształty i rozmiary. Zwykle jest on długi. Niekiedy występuje ogon chwytny, jak u badylarki pospolitej. Pokrywające ogon futro może być bujne – gryzonie żyjące na drzewach zazwyczaj mają puszysty ogon, biorący udział w utrzymaniu równowagi – może też być on kompletnie łysy.
Czasami służy on komunikacji. Bóbr uderza ogonem o wodę, a mysz domowa nim grzechocze, by wszcząć alarm. U niektórych gatunków istnieje jedynie szczątkowy ogon albo też nie mają go wcale, dotyczy to zwłaszcza form żyjących pod ziemią.
U niektórych gatunków istnieje możliwość regeneracji, jeśli część ogona się odłamie. Autotomię obserwuje się u myszarek i popielicowatych. Występuje u nich ulegająca łatwemu zsuwaniu się pochwa skórna dystalnego fragmentu ogona. Jej zsunięcie wiąże się z niewielkim tylko krwawieniem, po czym część ogona odpada. Nie następuje regeneracja.
Skóra
Skórę gryzoni pokrywa futro, ale niekiedy można spotkać u nich również kolce i łuski. Te ostatnie obserwuje się na ogonie myszowatych czy bobra. Istnieją też gryzonie o skórze nagiej, czego przykład stanowi golec piaskowy.
Gruczoły potowe nie są częste, w przeciwieństwie do gruczołów łojowych, często zgrupowanych.
Samice mają kilka par sutków (maksymalnie 11), rozlokowanych od okolicy piersiowej aż do pachwin. Nietypowo umiejscawiają się u nutrii – na grzbiecie – co ma związek z częściowo wodnym trybem życia tego ssaka.
Układ nerwowy
Mózgowie gryzoni zawiera dwie półkule mózgu, które zazwyczaj cechują się gładką powierzchnią, pofałdowanie zauważa się u większych przedstawicieli. Zwraca uwagę dobre rozwinięcie płatów węchowych.
Układ pokarmowy
Budowa układu pokarmowego gryzoni, zazwyczaj długiego, zależy od rodzaju spożywanego przez nie pokarmu (zazwyczaj jest to pokarm roślinny).
Żołądek gryzoni nie ma skomplikowanej budowy. Niekiedy jednak występuje w nim przewężenie pozwalające podzielić go na proksymalną część wpustową i dystalną część odźwiernikową. Anatomię taką obserwuje się u myszowatych.
Jelito ślepe występuje u gryzoni innych, niż popielicowate.
Układ rozrodczy
Jądra mogą leżeć w jamie brzusznej lub w pachwinie. Schowane w jamie brzusznej przez pozostałą część roku, na okres rozrodu przemieszczają się pod skórę brzucha bądź do moszny. Powiększają się one także w tym okresie. U wielu gatunków prącie zawiera w sobie kość zwaną baculum. Mogą je także zdobić kolce. U samców występują też dobrze rozwinięte dodatkowe gruczoły dróg płciowych.
Dymorfizm płciowy
Dymorfizm płciowy występuje u wielu gatunków. Niekiedy samce przerastają rozmiarami samice, kiedy indziej zdarza się sytuacja odwrotna. Przewaga samców typowa jest u świstaków, szczuroskoczków, gofferowatych. Prawdopodobnie rozwija się na skutek doboru płciowego i większych walk pomiędzy samcami. Przewaga samic występuje u gryzoni z rodzaju pręgowiec i u skoczkomyszek. Nie wiadomo, dlaczego się pojawia. W przypadku pręgowca sosnowego być może to samce wybierają sobie większe partnerki z uwagi na osiągany przez nie większy sukces rozrodczy. U niektórych gatunków, np. z podrodziny karczowników, dymorfizm płciowy zmienia się z populacji na populację. U nornicy rudej samice typowo przerastają samce, jednak większe rozmiary samców pojawiają się w populacjach alpejskich, prawdopodobnie z uwagi na brak drapieżników i większe współzawodnictwo pomiędzy samcami.
Rozmieszczenie geograficzne i siedlisko

Jako jedną z najbardziej rozpowszechnionych grup ssaków gryzonie spotyka się na każdym kontynencie z wyjątkiem Antarktyki. Jako jedyne lądowe łożyskowce skolonizowały Australię i Nową Gwineę bez udziału człowieka. Ludzie umożliwili tym stworzeniom rozprzestrzenienie się na wiele odizolowanych wysp oceanicznych. Przykład stanowi szczur polinezyjski. Rodentia przystosowały się do prawie wszystkich siedlisk lądowych, od mroźnej tundry (gdzie potrafią przeżyć pod śniegiem) do gorących pustyń. Niektóre zamieszkują drzewa, jak pewne wiewiórkowate czy ursonowate. Inne, jak gofferowate czy golec piaskowy, prowadzą prawie całkowicie podziemny tryb życia w zbudowanych przez siebie skomplikowanych systemach norw. Inne bytują na powierzchni ziemi, ale korzystają też z nory, w której mogą się schronić. Bobry i piżmak amerykański znane są z częściowo wodnego trybu życia, jednakże gryzoniem, który najlepiej zaadaptował się do wodnego środowiska, jest prawdopodobnie Crossomys moncktoni z Nowej Gwinei. Gryzonie radzą sobie także w siedliskach stworzonych przez człowieka: na obszarach rolniczych i miejskich.

Niektóre gatunki stanowią zwierzęta domowe. Istnieją też takie, które odgrywają istotną rolę w ekologii. Niektóre uznaje się za gatunki kluczowe, prowadzące inżynierię środowiskową w swych siedliskach. Na Wielkich Równinach Ameryki Północnej drążenie nor przez pieski preriowe odgrywa istotną rolę w napowietrzaniu gleby i redystrybucji składników odżywczych, w podnoszeniu zawartości substancji organicznych w glebie i zwiększaniu absorpcji wody. Nieświszczuki płowe utrzymują w ten sposób swe trawiaste siedliska. Pewni więksi roślinożercy, jak bizon amerykański i widłoróg, wolą paść się w pobliżu kolonii nieświszczuków płowych z uwagi na zwiększoną wartość odżywczą pokarmu. Pieski preriowe mogą również przyczyniać się do regionalnej czy lokalnej utraty bioróżnorodności, zwiększonego wyjadania ziarna oraz powstawania i rozprzestrzeniania się inwazyjnych krzewów. Gryzonie drążące w podłożu mogą zjadać owocujące grzybnie i rozpowszechniać zarodniki z wypróżnianym kałem, umożliwiając przez to grzybom rozprzestrzenianie i wchodzenie w mikoryzę z korzeniami roślin (które zazwyczaj wymagają tej symbiozy). W takiej sytuacji gryzonie odgrywają rolę w utrzymaniu zdrowego lasu.
W wielu regionach klimatu umiarkowanego bobry odgrywają kluczową rolę w hydrologii. Budując swe tamy i żeremie, ssaki te modyfikują przebieg strumieni i rzek, umożliwiając tworzenie rozległych siedlisk wodnych. Jedno z badań wykazało, że inżynieria bobrów prowadzi do wzrostu o 33% liczby gatunków roślin zielnych w strefie przybrzeżnej. Inne stwierdziło, że bobry zwiększają liczebność populacji dzikiego łososia.
Zachowanie
Pożywienie

Większość gryzoni to roślinożercy, żywiący się wyłącznie pokarmem roślinnym, jak nasiona, liście, kwiaty i korzenie. Niektóre są jednak wszystkożerne, a zdarzają się nieliczne drapieżniki. Nornik bury to typowy roślinożerny gryzoń. Pasie się na trawie, ziołach, bulwach, mchach i innej roślinności. W zimie podgryza korę. Okazjonalnie spożywa bezkręgowce, jak larwy owadów. Goffer równinny zjada materiał roślinny znajdowany pod ziemią w czasie drążenia tuneli, zbiera też trawy, korzenie i bulwy w swych workach policzkowych, by przenieść je pod ziemię do swej spiżarni. Jego krewny, goffer brunatny (Geomys personatus) unika wychodzenia na powierzchnię w celu poszukiwania pokarmu. Żywi się, chwytając pyskiem korzenie roślin i ciągnąc je w dół do swej nory. Praktykuje również koprofagię. Wielkoszczur leśny (Cricetomys emini) żeruje na powierzchni, gromadząc wszystko, co tylko może być jadalne, w swych pojemnych workach policzkowych. Na jego twarzy pokazują się wybrzuszenia kierujące się bocznie. Wtedy dopiero wraca do swej nory posortować zgromadzony materiał i zjeść to, co się nadaje do zjedzenia.
Aguti należy do nielicznych zwierząt potrafiących dobrać się do dużych torebek orzesznicy wyniosłej. W środku znajduje się jednak zbyt wiele nasion, by zwierzę spożyło je na jeden raz, w związku z czym aguti zabiera część ze sobą. Wspomaga to rozprzestrzenianie się nasion. Z tych niewykorzystanych przez aguti wyrosną rośliny, które wykiełkują z dala od organizmów rodzicielskich. Inne drzewa rodzące orzechy przejawiają tendencję do produkowania ich jesienią w nadmiarze. Owoce występują wtedy zbyt licznie, by spożyć je jednorazowo. Wiewiórki gromadzą i magazynują je w szczelinach bądź dziuplach drzew. W regionach pustynnych nasiona dostępne są często tylko przez krótki okres. Szczuroskoczki zbierają wszystkie, jakie znajdą, a następnie składują w spiżarni swojej nory.

Strategia radzenia sobie z sezonową obfitością pokarmu polega na jedzeniu, ile tylko się da i magazynowaniu nadmiaru w postaci tkanki tłuszczowej. Występuje ona u świstaków, które to gryzonie mogą być jesienią o 50% cięższe niż wiosną. Podczas długiej zimy zwierzęta korzystają z nagromadzonych rezerw tłuszczu, ulegając hibernacji. Bobry żywią się liśćmi, pączkami i wewnętrzną warstwą kory wzrastających drzew, jak też roślinami wodnymi. Magazynują żywność na zimę, powalając niewielkie drzewa i ulistnione gałęzie jesienią, zanurzając je pod wodą i zagrzebując koniec w mule, by je zakotwiczyć. Dzięki temu mają dostęp do zapasów żywności przetrzymywanych pod wodą, nawet kiedy lód pokryje staw.
Choć tradycyjnie uznaje się je za roślinożerców, liczne gatunki oportunistycznie zjadają owady, ryby, mięso, a bardziej wyspecjalizowane formy polegają na takim pokarmie. Badanie funkcjonalno-morfologiczne uzębienia gryzoni wskazuje na to, że prymitywne gryzonie były raczej wszystkożerne niż roślinożerne. Przeglądy literatury ukazują, że liczni członkowie Sciuromorpha i Myomorpha, a także kilku przedstawicieli Hystricomorpha włączają do swojej diety pokarm zwierzęcy lub też spożywają taki pokarm, gdy zaoferuje im się go w niewoli. Badanie zawartości żołądków żyjącego na kontynencie północnoamerykańskim myszaka białostopego (Peromyscus leucopus), uznawanego normalnie za roślinożercę, wykazało 34% materii zwierzęcej.
Bardziej wyspecjalizowani mięsożercy to ryjoszczury z Filipin, żywiące się owadami i bezkręgowcami o miękkim ciele, australijskie bobroszczury, żywiące się owadami wodnymi, rybami, skorupiakami, małżami, ślimakami, płazami bezogonowymi, jajami i ptakami wodnymi. Pasikoniszka białobrzucha, zamieszkująca suche regiony Ameryki Północnej, żywi się owadami, skorpionami, innymi małymi myszami, a jedynie niewielką część jej diety stanowi pokarm roślinny. Ma krępe ciało z krótkimi łapami i ogonem, ale jest zwinna i potrafi z łatwością pokonać zdobycz swojej wielkości.
Zachowania społeczne

Gryzonie wykazują szeroki zakres zachowań społecznych. Występuje wśród nich pierwszy poznany system kastowy wśród ssaków, spotykany u golca piaskowego. U pieska preriowego występują rozległe kolonie. Dalej spotyka się grupy społeczne, a także samotniczy tryb życia u popielicy szarej, której dorosłe osobniki mają zachodzące na siebie areały żerowania, żyją jednak w osobnych gniazdach i żerują samodzielnie. Gromadzą się na krótko podczas sezonu rozrodczego w celu dobierania się w pary. Goffer również żyje samotnie poza sezonem rozrodczym. Każdy osobnik drąży złożony układ tuneli i utrzymuje własne terytorium.
Większe gryzonie wykazują tendencję do życia w grupach rodzinnych, w których rodzice i dzieci żyją razem, nim potomstwo odejdzie. Bobry żyją w rozszerzonych jednostkach rodzinnych obejmujących typowo parę dorosłych, tegoroczne młode, ich rodzeństwo z poprzedniego roku oraz czasami jeszcze starsze młode. Szczur wędrowny zazwyczaj tworzy niewielkie kolonie. Do sześciu samic dzieli wspólną norę, a pojedynczy samiec broni terytorium wokół niej. W przypadku dużego zagęszczenia populacji system ten załamuje się i wśród samców ustala się system hierarchiczny oparty na dominacji i zazębiających się areałach. Córki pozostają w kolonii, podczas gdy synowie opuszczają ją. Nornik preriowy (Microtus ochrogaster) jest monogamiczny, tworzy długo utrzymujące się pary. Poza sezonem rozrodu żyje w bliskim sąsiedztwie innych osobników w obrębie małej kolonii. Samiec nie wykazuje agresji w kierunku innych samców, póki nie paruje się z samicą. W tym czasie broni swego terytorium, samicy i gniazda przed innymi samcami. Para tuli się do siebie, czyści się wzajemnie, gniazduje razem i dzieli się odpowiedzialnością za potomstwo.

Do najbardziej społecznych gryzoni należą świstaki. Typowo tworzą one kolonie opierające się na więziach pomiędzy samicami. Samce po odstawieniu od sutka rozchodzą się i jako osobniki dorosłe wiodą wędrowny tryb życia. Współpraca u susłów i świszczy różni się pomiędzy gatunkami. Zazwyczaj obejmuje alarmowanie się przez zagrożeniem, obronę terytorium, dzielenie się pokarmem, ochronę miejsc gniazdowania i przeciwdziałanie dzieciobójstwie. Nieświszczuk płowy buduje wielkie miasta, mogące rozciągać się na wiele hektarów. Jamy nie łączą się ze sobą. Są drążone i zamieszkiwane przez terytorialne grupy rodzinne zwane koteriami. Koteria zwykle składa się z dorosłego samca, trzech lub czterech dorosłych samic, kilku nie przystępujących do rozrodu roczników i młodych z ostatniego roku. Osobniki w obrębie koterii są do siebie przyjaźnie nastawione, wrogo natomiast do innych.
Prawdopodobnie najbardziej ekstremalny przykład zachowań kolonialnych obserwuje się u golca piaskowego i zęboszczura kolonijnego, gryzoni eusocjalnych. Golce całe swe życie spędzają pod ziemią. Tworzą kolonie, które mogą sięgać 80 osobników. Do rozrodu przystępuje w nich jedna samica i do trzech samców. Inni członkowie kolonii osiągają mniejsze rozmiary, są bezpłodni i funkcjonują w grupie jako robotnicy. Pewne osobniki mają rozmiary pośrednie. Pomagają oni w odchowie młodych. W razie śmierci jednego z osobników rozmnażających się mogą zająć jego miejsce. U zęboszczura kolonijnego w jednej kolonii występuje jeden aktywnie seksualnie samiec i samica, podczas gdy pozostałe osobniki nie są właściwie bezpłodne, ale mogą stać się płodne tylko wtedy, gdy założą własną kolonię.
Komunikacja
Węch

Gryzonie wykorzystują zapachy w wielu sytuacjach, w tym w komunikacji wewnątrz- i międzygatunkowej, oznaczaniu ścieżek i zakładaniu terytoriów. Mocz dostarcza informacji genetycznych, w tym o gatunku, płci i konkretnym osobniku, a także informacji metabolicznych na temat dominacji, zdrowia i statusu reprodukcyjnego. Związki związane z głównym układem zgodności tkankowej (MHC) wiążą się z kilkoma białkami moczu. Zapach drapieżnika hamuje jednak epatowanie własnym.
Gryzonie potrafią rozpoznawać swych bliskich krewnych węchem. Pozwala im to wyróżniać swych krewnych, a także unikać chowu wsobnego. Członków rodziny poznają po zapachu moczu, kału i wydzielin gruczołów. Główną rolę może tu odgrywać MHC, gdyż stopień spokrewnienia dwóch osobników koreluje z liczbą wspólnych genów MHC. W komunikacji z osobnikami niespokrewnionymi, w przypadku której zachodzi potrzeba użycia bardziej trwałych markerów zapachowych, choćby przy oznaczaniu granic terytorium, wykorzystywane są nielotne białka moczu (major urinary proteins, MUP), działające jak transportery feromonów. MUP także mogą wskazywać na osobnika. Każdy samiec myszy domowej wydala mocz zawierający około tuzina zależnych od jego genomu MUP.
Mysz domowa znaczy terytorium moczem, który zawiera feromony. Podobnie wykorzystuje mocz w rozpoznawaniu się osobników i kontaktach społecznych. Może to czynić na różne sposoby
- efekt Bruce: feromony obcych samców wywołują zakończenie ciąży samic
- efekt Whittena: feromony znajomych samców wywołują u populacji samic zsynchronizowaną ruję
- efekt Vandenbergha: feromony dojrzałego samca myszy domowej powodują wczesną indukcję pierwszego cyklu rujowego u prawie dojrzałej samicy
- efekt Lee–Boota: feromony dojrzałej samicy wywołują supresję lub wydłużenie cyklu rujowego innej samicy przebywającej z nią w jednej grupie, w której nie ma samców
- feromony samca bądź ciężarnej lub karmiącej samicy mogą przyśpieszać bądź opóźniać dojrzewanie płciowe młodocianych samic.
Terytorialne bobry czy wiewiórki (sosnowiórka czerwona) poszukują i zaznajamiają się z zapachami swych sąsiadów. Odpowiadają mniej agresywnie, jeśli to któryś z nich dokona wtargnięcia, niż gdyby to był osobnik obcy czy poszukujący własnego terytorium. Po angielsku określa się to zjawisko terminem „dear enemy effect”.
Słuch

Wiele gatunków gryzoni, zwłaszcza wiodących dzienny i społeczny tryb życia, dysponuje szerokim zakresem dźwięków alarmowych, wydawanych w razie dostrzeżenia niebezpieczeństwa. Z takiego zachowania wynikają korzyści, tak pośrednie, jak i bezpośrednie. Potencjalny drapieżnik może zatrzymać się, kiedy zorientuje się, że odkryto jego obecność. Odgłos ostrzegawczy pozwala innym osobnikom tego samego gatunku, w tym krewnym, podjąć odpowiednie działania. Kilka gatunków, jak choćby nieświszczuk płowy, posługuje się złożonymi systemami dźwięków ostrzegających przed drapieżnikami. Zwierzęta te mogą wydawać z siebie różne odgłosy w zależności od tego, z jakim drapieżnikiem mają do czynienia (np. czy chodzi o drapieżcę chodzącego po ziemi, czy atakującego z powietrza). Każdy dźwięk zawiera w sobie informacje o naturze zagrożenia. Nagłość niebezpieczeństwa komunikowana jest przez cechy akustyczne odgłosów.
Gryzonie prowadzące społeczny tryb życia dysponują szerszym repertuarem dźwięków, niż gatunki samotnicze. U dorosłych przedstawicieli zęboszczurów rozpoznano 15 różnych odgłosów, u młodocianych zaś 4. Podobnie żyjąca w społeczności koszatniczka pospolita, gryzoń kopiący nory, wykazuje szeroki wachlarz metod komunikacji i posługuje się również szerokim repertuarem dźwięków, obejmującym 15 ich różnych rodzajów. Popielicowate porozumiewają się ze sobą z użyciem ultradźwięków, gdy nie widzą się wzajemnie. Mysz domowa również słyszy ultradźwięki, tak jak dźwięki słyszalne dla człowieka. Wykorzystuje je w różnych sytuacjach. Odgłosy słyszane przez ludzi padają często podczas przyjacielskich bądź wrogich spotkań. Ultradźwięki wykorzystywane są w celach rozrodczych oraz przez młode, które wypadły z gniazda.
Szczury laboratoryjne, zaliczane do gatunku szczur wędrowny, wydają z siebie krótkie ultradźwięki o wysokiej częstotliwości podczas rzekomo przyjemnych sytuacji, jak zabawa w zapasy, gdy antycypują podaż morfiny, podczas parzenia się czy łaskotania. Wokalizacje, określane jako świergotanie, porównywano do śmiechu i interpretowano jako coś pozytywnego. W badaniach klinicznych świergot wiąże się z pozytywnymi emocjami, wytwarza się więź z osobą łaskoczącą. W efekcie badane szczury szukały łaskotania. Jednak z wiekiem tendencja do świergotania spada. Jak większość odgłosów szczurów, świergot wydawany jest z częstotliwością zbyt dużą dla ludzkiego ucha niewyposażonego w specjalistyczny sprzęt. By stwierdzić świergot, wykorzystuje się urządzenie służące do wykrywania nietoperzy.
Odnotowano, że szczury wędrowne wykorzystują ultradźwięki w echolokacji. Częstotliwości odbierane przez uszy gryzoni różnią się pomiędzy gatunkami. Wybrane przykłady obrazuje poniższa tabela.
| Takson | Dolna granica (Hz) | Górna granica (Hz) | Specyficzne odgłosy |
|---|---|---|---|
| człowiek (dla porównania) | 64 | 23.000 | |
| szczur | 200 | 76.000 | ultradźwięki wysokiej częstotliwości (50 kHz) w razie przyjemności |
| mysz | 1.000 | 91.000 | młode poddane stresowi wydaje dźwięki o częstotliwości 40 kHz |
| myszoskoczki | 100 | 60.000 | |
| kawia domowa (świnka morska) | 54 | 50.000 | |
| szynszyla | 90 | 22.800 |
Wzrok
Gryzonie, jak wszystkie łożyskowce z wyjątkiem naczelnych, dysponują jedynie dwoma rodzajami czopków siatkówki. Odbierają one światło niebieskie-UV o krótkiej fali oraz zielone o fali pośredniej. Wobec tego są dichromatyczne. Dostrzegają jednak ultrafiolet, widzą więc światło, którego nie zauważa człowiek. Nie wiadomo do końca, jaką funkcję pełni dostrzeganie przez nie UV. Przykładowo brzuch koszatniczki odbija więcej światła UV, niż jej grzbiet. Jeśli więc zwierzę to, zaniepokojone, wstanie na tylne łapy, inne osobniki zobaczą jego brzuszną stronę ciała, co może je zaalarmować. Jeśli gryzoń stoi na czterech łapach, jego gorzej odbijający promienie UV grzbiet czyni go mniej widocznym dla drapieżników. Za dnia światło ultrafioletowe dostępne jest w obfitości, jednak w nocy już nie. Rankiem dochodzi do znacznego wzrostu stosunku natężenia światła ultrafioletowego do widzialnego, odwrotnie wieczorem. Wiele gryzoni wykazuje aktywność zmierzchową. Wrażliwość na UV byłoby więc dla nich znacznym udogodnieniem. Nie wiadomo, jaką wartość miałoby widzenie ultrafioletu dla gatunków nocnych.
Mocz wielu gryzoni (wymienia się tu norniki, koszatniczkę, myszy, szczury) silnie odbija światło UV, co może odgrywać rolę w komunikacji, podobnie jak sygnały zapachowe. Jednak liczba odbijanych promieni UV spada z czasem, co w pewnych warunkach niesie za sobą niekorzystny efekt. Pustułka zwyczajna potrafi odróżnić stare i świeże ślady obecności gryzoni, dzięki polowaniu nad jedynie świeżo oznaczonymi ścieżkami odnosi większy sukces w łowiectwie.
Dotyk

Wibracje mogą dostarczyć innym osobnikom tego samego gatunku informacji na temat podejmowanych zachowań, niebezpieczeństwa ze strony drapieżnika, utrzymania grupy oraz zalotów. Ślepiec lewantyński był pierwszym ssakiem, w przypadku którego udokumentowano porozumiewanie się za pomocą wywoływania wibracji podłoża. Ten drążący pod ziemią gryzoń uderza głową w ściany budowanych przez się korytarzy. Pierwotnie uważano, że zachowanie to stanowi część budowania przezeń tuneli, jednakże później odkryto, że tworzy ono uporządkowane w czasie sygnały docierające na dalekie odległości do sąsiadujących osobników
Tupanie jest często wykorzystywane w celu ostrzeżenia przed drapieżnikiem bądź w obronie. Stosują je głównie gryzonie żyjące w całości lub po części od ziemią. Szczuroskoczek flagoogonowy wytwarza kilka wzorców tupania stosowanych w różnych sytuacjach, choćby w razie napotkania węża. Tupanie może ostrzec pobliskie potomstwo, ale najbardziej prawdopodobne wyjaśnienie podaje, że szczuroskoczek jest zbyt czujny na uwieńczony sukcesem atak i dlatego zauropsyd go nie goni. Kilka badań wskazywało na celowe wykorzystanie wibracji podłoża do śródgatunkowej komunikacji u piaskogrzeba przylądkowego. Tupanie wchodzi w skład rywalizacji pomiędzy samcami. Osobnik dominujący zaznacza poprzez dudnienie swój potencjał i stan posiadania, minimalizując dzięki temu ryzyko kontaktu z potencjalnym rywalem.
Rozmnażanie
Systemy dobierania się

Niektóre gatunki gryzoni wiodą monogamiczne życie. Dorosły samiec i dorosła samica tworzą u nich długo trwający związek. Monogamia u gryzoni może przybierać dwie formy: obligatoryjną i fakultatywną. W pierwszym przypadku oboje rodziców sprawuje opiekę nad potomstwem i odgrywa istotną rolę w zwiększeniu szans na przetrwanie swych dzieci. Jest tak u myszaka kalifornijskiego, myszaka plażowego, malgaszoszczura wielkiego i bobrów. Samce w przypadku takich gatunków kopulują jedynie ze swymi partnerkami. Poza zwiększoną opieką nad potomstwem obligatoryjna monogamia zmniejsza ryzyko, że samiec nigdy nie znajdzie partnerki bądź pokryje bezpłodną samicę. W monogamii fakultatywnej samiec nie sprawuje bezpośredniej opieki rodzicielskiej. Zostaje z jedną samicą, ponieważ nie ma szans na znalezienie innej z powodu przestrzennego rozmieszczenia osobników. Przykład wydaje się stanowić tutaj nornik preriowy (Microtus ochrogaster). Samce tego gatunku strzegą i bronią samic w swym sąsiedztwie.
Samce gatunków poligamicznych starają się zmonopolizować dostęp do wielu samic i kopulować z nimi. Podobnie jak w przypadku monogamii, poligamia może przybierać u gryzoni dwie formy: obronną i nieobronną. W przypadku tej pierwszej samiec kontroluje terytorium zawierające zasoby, które przywabiają płeć przeciwną. Dzieje się tak u świstaków, np. u świstaka żółtobrzuchego, susłoucha plamkowanego, susłogona rudoczelnego i susłogona preriowego. Samce panują więc na własnych terytoriach jako ich „rezydenci” Na ich terytoriach żyją też samice, „rezydentki”. W przypadku świstaków wydaje się, że samce nigdy nie tracą swych rewirów i zawsze wygrywają w potyczkach z samcami-intruzami. W przypadku pewnych gatunków bezpośrednio broniących swych samic występują walki skutkujące nieraz poważnymi ranami. Jeśli chodzi o poligamię nieobronną, samce nie wykazują terytorializmu i podróżują w poszukiwaniu samic, by je posiąść. Pomiędzy nimi wytwarza się hierarchia ustalająca, które osobniki nad którymi dominują. Osobniki stojące na szczycie zyskują dostęp do większości samic. Taki stan rzeczy obserwuje się u susłogona łakowego i pewnych wiewiórek drzewnych.
Promiskuityzm polega na posiadaniu licznych partnerów seksualnych przez zarówno samce i samice. Również spotyka się go u gryzoni. U gatunków takich jak myszak białostopy samice wydają na świat mioty, w których poszczególnych młodych nie łączy wspólne ojcostwo. Promiskuityzm prowadzi do zwiększonej konkurencji pomiędzy nasieniem różnych samców, które mają w związku z tym większe jądra. Jądra afrowiórki namibijskiej mogą osiągać 20% długości jej głowy i tułowia. Pewne gatunki gryzoni rozwinęły elastyczne systemy parzenia się, wahające się pomiędzy monogamią, poligamią i promiskuityzmem.
Samice odgrywają istotną rolę w doborze sobie samca. W swych wyborach biorą pod uwagę wielkość, pozycję w hierarchii i terytorium samca. U eusocjalnych golców pojedyncza samica monopolizuje rozród z przynajmniej trzema samcami.
U większości gatunków, jak szczur wędrowny czy mysz domowa, owulacja następuje w regularnym cyklu. U innych, jak norniki, indukuje ją kopulacja. W trakcie tej ostatniej samce niektórych gatunków pozostawiają mating plug w pochwie samicy. Spełnia on dwojaką rolę: zabezpiecza nasienie przed wypłynięciem na zewnątrz, a samicę przed zaplemnieniem przez innego samca. Samica potrafi jednak usunąć ten czop, zarówno zaraz po połączeniu, jak i po kilku godzinach.
Narodziny i opieka nad potomstwem

Gryzonie mogą rodzić się gniazdownikami (ślepe, nieowłosione, względnie niedorozwinięte) bądź zagniazdownikami (w większości owłosione, z otwartymi oczami, dobrze rozwinięte). Zależy to od gatunku. Wiewiórkowate czy myszowate należą do tej pierwszej grupy. Do zagniazdowników zaś zaliczają się kawie domowe i jeżozwierzowate. Matki gniazdowników zazwyczaj budują wypracowane gniazda, nim wydadzą swe potomstwo na świat. Utrzymują je aż do odstawienia swych młodych. Samica rodzi na siedząco lub na leżąco. Noworodki wydostają się w kierunku, w który zwraca się matka. Młode opuszczają gniazdo po raz pierwszy kilka dni po tym, jak otwierają oczy i początkowo regularnie wracają do domu. Gdy z czasem stają się coraz lepiej rozwinięte, odwiedzają gniazdo coraz rzadziej i po odstawieniu opuszczają je na zawsze.
Matka zagniazdowników mało się stara przy budowie gniazda, niekiedy w ogóle nie zawraca sobie tym głowy. Rodzi na stojąco i młode wydostają się na świat za nią. Utrzymuje kontakt głosowy ze swymi wysoce ruchliwymi dziećmi. Są one względnie niezależne i odstawienie następuje po kilku dniach. Jednak opieka matki (karmienie, czyszczenie) trwa nadal. Miot gryzoni może przybierać różną liczebność. Samice, które wydały na świat mniej liczne mioty, mniej czasu spędzają w gnieździe.

Matka sprawuje bezpośrednią opiekę nad swym potomstwem. Karmi je, czyści, przenosi i tuli. Zajmuje się też nim pośrednio, tzn. zapewnia mu pożywienie, gniazdo i ochronę. U wielu gatunków społecznych opiekę nad młodymi może również sprawować osobnik inny, niż ich rodzic. Sytuację tę określają anglojęzyczne terminy alloparenting lub cooperative breeding. Sytuacja taka ma miejsce u nieświszczuka płowego i susłogona łąkowego, u których matki żyją we wspólnych gniazdach i karmią cudze młode wraz ze swoimi. Nasuwa się pytanie, czy samice te potrafią w ogóle odróżnić własne młode od cudzych. Młode mary patagońskie także żyją we wspólnych siedzibach, jednak matki nie pozwalają nieswoim młodym ssać.
Wśród licznych gatunków gryzoni dochodzi do dzieciobójstwa. Praktykują je osobniki dorosłe obu płci tego samego gatunku co młode. Zaproponowano kilka możliwych wyjaśnień tego zachowania, wśród nich stres związany z ilością pożywienia, konkurencję o zasoby, unikanie niewłaściwie ukierunkowanej opieki nad potomstwem i, jeśli chodzi o samce, czynienie matki na powrót gotową na przyjęcie adoratora. To ostatnie postępowanie jest dobrze znane u naczelnych i lwów, gorzej natomiast znane u gryzoni. Dzieciobójstwo zdaje się być szeroko rozpowszechnione u nieświszczuka płowego, włączając w to zabójstwa dokonywane przez wkraczające na cudzy teren samce i imigrujące samice, jak też okazjonalny kanibalizm – pożeranie własnego potomstwa. Dla ochrony przez dzieciobójstwem ze strony innych osobników dorosłych samice unikają ich bądź też okazują bezpośrednią agresję potencjalnym sprawcom, łączą się z licznymi samcami, występuje wśród nich terytorializm i wczesne rozwiązywanie ciąży. U niektórych gryzoni obserwuje się też zabijanie płodów. Samica dominująca świstaka stara się powstrzymać reprodukcję samic podporządkowanych. Zachowuje się w stosunku do nich wrogo, jeśli zajdą w ciążę. Stresuje je, co powoduje u nich poronienie.
Inteligencja

Gryzonie cechują się zaawansowanymi zdolnościami poznawczymi. Potrafią szybko uczyć się unikać zatrutych przynęt, co czyni je trudnymi do pozbycia się szkodnikami. Kawia domowa potrafi nauczyć się i zapamiętać skomplikowane ścieżki prowadzące do jedzenia. Wiewiórki i szczuroskoczki potrafią lokalizować pokarm dzięki pamięci przestrzennej raczej niż za pomocą węchu.
Ponieważ laboratoryjne myszy domowe i szczury wędrowne są szeroko wykorzystywane jako modele w badaniach naukowych, nastąpił duży postęp w wiedzy o ich zdolnościach poznawczych. Szczury wędrowne wykazują błąd poznawczy, obróbka informacji zależy u nich od pozytywnego bądź negatywnego afektu. Przykładowo szczury laboratoryjne trenowane do odpowiedzi na specyficzny ton poprzez naciśnięcie dźwigni i otrzymanie nagrody, a innej dźwigni w odpowiedzi na inny ton w celu uniknięcia kary w postaci rażenia prądem, z większym prawdopodobieństwem odpowiedzą na ton pośredni, naciskając dźwignię związaną z nagrodą, jeśli przed chwilą ktoś je połaskotał (co uwielbiają).
Laboratoryjne szczury wędrowne mogą mieć zdolność metapoznania – rozważania swego własnego uczenia i podejmowania decyzji na podstawie tego, co wiedzą lub czego nie wiedzą, co wskazywały podejmowane przez nie wybory w związku z poziomem trudności zadań i oczekiwaną nagrodą. Byłyby więc szczury pierwszymi po naczelnych zwierzętami, u których wykryto taką zdolność. Jednakże wyniki te są dyskusyjne. Za zachowanie szczurów odpowiadać mogło po prostu warunkowanie instrumentalne lub model ekonomii behawioralnej.
Szczury wędrowne wykorzystują społeczne uczenie się w szerokim zakresie sytuacji, szczególnie jednak jeśli chodzi o preferencje zdobywania pożywienia. Uczenie się o odpowiednich zasobach pożywienia podzielić można na 4 etapy:
- przed urodzeniem, w macicy płód wykrywa cząsteczki przenoszące zapach pochodzące z pokarmu jego matki, przenikające barierę łożyskową. Krótko po urodzeniu noworodek odpowiada pozytywnie na pokarmy, które wyczuwał in utero.
- w czasie karmienia przez matkę młode uzyskuje informację o diecie swej matki zawartą w mleku. Preferuje później pokarm, który matka jadała podczas laktacji.
- gdy młode szczury są odstawiane od piersi i zaczynają spożywać stały pokarm, osobniki dorosłe służą im za przewodników. Żerują tam, gdzie żerują dorosłe zwierzęta lub gdzie te ostatnie pozostawiły swój zapach.
- w okresie adolescencji i dorosłego życia wybór pożywienia zależy od interakcji społecznych i może zachodzić daleko od miejsc żerowania. Szczury wyczuwają zapachy na futrze, wibryssach i w oddechu innych osobników. Silnie preferują jedzenie, które wcześniej spożywały inne szczury. Oznacza to, że nie będą jeść trutek unikanych przez starsze osobniki.
Ewolucja

Gryzonie w zapisie kopalnym rozpoznaje się głównie po uzębieniu. Pojawiają się w nim w paleocenie, niedługo po wymieraniu kredowym około 66 milionów lat temu. Skamieliny te pochodzą z Laurazji, superkontynentu zbudowanego z dzisiejszych lądów Ameryk Północnej, Europy i Azji. Siekaczowce (Glires), klad obejmujący gryzonie i zajęczaki, oddzielił się od innych łożyskowców w kenozoiku, radiacja ewolucyjna nastąpiła w okolicy granicy kreda-paleogen. Dane uzyskane z wykorzystaniem zegara molekularnego sugerują, że rząd Rodentia pojawił się już w kredzie późnej, ale inne szacunki również oparte na danych molekularnych pozostają w zgodzie z zapisem kopalnym.
Historia kolonizacji kontynentów przez gryzonie jest złożona. Pochód wielkiej nadrodziny myszowych (Muroidea) (chomikowate, myszowate) może obejmować aż do 7 kolonizacji Afryki, 5 – Ameryki Północnej, 4 – Azji Południowo-Wschodniej, 2 – Ameryki Południowej i do 10 – Eurazji.

W eocenie gryzonie zaczęły się różnicować. Bobry pojawiły się w Ameryce Północnej w późnym eocenie, nim rozprzestrzeniły się na Eurazję. Późno w eocenie jeżozwierzokształtne (Hystricognathi) skolonizowały Afrykę, powstając najprawdopodobniej w Azji przynajmniej 39,5 miliona lat wcześniej. Zapis kopalny obrazuje dalej migrację z Afryki na kontynent południowoamerykański, wcześniej izolowany, prawdopodobnie z wykorzystaniem prądów oceanicznych, dzięki którym gryzonie przebyły Ocean Atlantycki na naturalnych tratwach. Caviomorpha przybyły do Ameryki Południowej 41 milionów lat temu, przynajmniej tak wcześnie, jak Hystricognatha do Afryki, osiągając Wielkie Antyle we wczesnym oligocenie. Sugeruje to, że musiały szybko rozprzestrzenić się w Ameryce Południowej.
20 milionów lat temu w zapisie kopalnym pojawiły się skamieniałości w widoczny sposób przynależące do współczesnych rodzin, takich jak myszowate. Do miocenu, kiedy to Afryka zderzyła się z Azją, afrykańskie gryzonie, jak jeżozwierze, rozprzestrzeniły się w Eurazji. Pewne gatunki kopalne osiągały bardzo duże rozmiary w porównaniu z dzisiejszymi ich krewnymi. Bóbr olbrzymi Castoroides ohioensis osiągał 2,5 m długości oraz 100 kg masy ciała. Jednak największym znanym gryzoniem jest Josephoartigasia monesi, przedstawiciel pakaranowatych, o długości ciała 3 m.
Pierwsze gryzonie przybyły do Australii przez Indonezję około 5 milionów lat temu. Na kontynencie tym najbardziej rozwiniętą grupą ssaków są torbacze, żyje tam też wiele gryzoni należących do podrodziny myszy (Murinae). Około 50 gatunków „starych” endemitów pochodzi od pierwszej fali, która pojawiła się tu w miocenie i wczesnym pliocenie. Spośród 10 gatunków szczurów 8 określa się jako „nowe endemity”. Ich przodkowie przybyli w kolejnej fali w późnym pliocenie lub wczesnym plejstocenie. Najwcześniejsze skamieniałości gryzoni z Australii mają maksymalnie 4,5 miliona lat. Dane molekularne potwierdzają, że po kolonizacji Nowej Gwinei podczas późnej epoki mioceńskiej i wczesnej plioceńskiej nastąpiło szybkie różnicowanie się. Kolejna fala radiacji adaptacyjnej po jednej bądź większej liczbie kolonizacji zaszła jakieś 2-3 miliony lat później. Dwa gatunki zostały niedawno introdukowane do Australii przez Europejczyków, nie należą więc do endemitów.
Po połączeniu się Ameryk Przesmykiem Panamskim około 3 miliony lat temu pewne gryzonie wzięły udział w wielkiej wymianie amerykańskiej. Nieznaczna liczba gatunków, jak pewne ursonowate, skierowała się na północ, natomiast bawełniaki (Sigmodontinae) ruszyły na południe. Zróżnicowały się tam szybko lub, wedle innego poglądu, zróżnicowały się już w Ameryce Północnej i Środkowej przed przekroczeniem nowego pomostu lądowego.
Klasyfikacja
Nazwę „Rodentia” (od łacińskiego rodere – gryźć) wprowadził angielski podróżnik i przyrodnik Thomas Edward Bowdich (1821). Niegdyś do rzędu włączano króliki, zające i szczekuszki, posiadające stale wzrastające jak u gryzoni siekacze. Posiadają one jednak dodatkową parę siekaczy w szczęce i grupuje się je w oddzielnym rzędzie zajęczaków. Te dwie grupy mają odmienną historię ewolucyjną. Filogeneza gryzoni umieszcza je w obrębie kladów Glires, Euarchontoglires i Boreoeutheria. Poniższy kladogram prezentuje zarówno zewnętrzne, jak i wewnętrzne pokrewieństwa gryzoni. Powstał dzięki pracy Wu et al., którzy uwzględnili zarówno dane paleontologiczne, jak i molekularne. Opublikowano go w 2012.
| Boreoeutheria |
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Rząd gryzoni (Rodentia) dzielony jest na podrzędy, infrarzędy, nadrodziny i rodziny. W tej grupie bardzo silnie zaznaczył się paralelizm ewolucyjny i konwergencja. Miały one bowiem tendencję do ewolucji w kierunku zajmowania z grubsza podobnych nisz ewolucyjnych. Ewolucja równoległa przejawia się nie tylko w budowie zębów, ale też podoczodołowego regionu czaszki. Utrudnia to klasyfikację, gdyż podobne cechy mogą nie wynikać wcale ze wspólnego pochodzenia. Brandt w 1855 po raz pierwszy zaproponował podział gryzoni na trzy podrzędy: Sciuromorpha (wiewiórkokształtne), Hystricomorpha (jeżozwierzowce) i Myomorpha (myszokształtne). Oparł ten pogląd na rozwoju pewnych mięśni szczęki. System ten powszechnie zaakceptowano. Schlosser w 1884 sporządził obszerny przegląd skamieniałości gryzoni. W większości użył doń zębów policzkowych. Zauważył, że pasują do klasycznej klasyfikacji. Jednak Tullborg w 1899 zaproponował wyróżnianie jedynie dwóch podrzędów: Sciurognathi i Hystricognathi. Autor bazował na wygięciu żuchwy. Dalej podzielił je na Sciuromorpha, Myomorpha, Hystricomorpha i Bathyergomorpha. Matthew w 1910 stworzył drzewo obrazujące filogenezę gryzoni Nowego Świata. Nie objął nim jednak problematycznych Rodentia Starego Świata. Dalsze wysiłki nie doprowadziły do zgody w tej kwestii. Niektórzy autorzy zaadaptowali klasyczny system z trzema podrzędami, inni dwa podrzędy Tullborga.
Niezgodności pozostawały bez rozwiązania. Także badania molekularne nie rozwiązały tej kwestii w pełni, choć potwierdziły one monofiletyzm gryzoni, a także pochodzenie kladu od wspólnego przodka żyjącego w paleocenie. Carleton i Musser (2005) w Mammal Species of the World prowizorycznie przyjęli system 5 podrzędów: Sciuromorpha, Castorimorpha (bobrokształtne), Myomorpha, Anomaluromorpha (wiewiórolotkowe) i Hystricomorpha. Występują w nim 33 rodziny, 481 rodzajów i 2277 gatunków.
Systamatyka na podstawie Illustrated Checklist of the Mammals of the World (2020):
Rząd gryzonie – Rodentia





- podrząd: Supramyomorpha
- infrarząd: Castorimorphi – bobrokształtne
- rodzina: Castoridae – bobrowate
- rodzina: Heteromyidae – karłomyszowate
- rodzina: Geomyidae – gofferowate
- infrarząd: Anomaluromorphi – wiewiórolotkowe
- rodzina: Zenkererllidae – nielotki
- rodzina: Anomaluridae – wiewiórolotkowate
- rodzina: Pedetidae – postrzałkowate
- infrarząd: Myomorphi – myszokształtne
- nadrodzina: Dipodoidea – skoczkowce
- rodzina: Sminthidae – smużki
- rodzina: Zapodidae – skoczomyszki
- rodzina: Dipodidae – skoczkowate
- nadrodzina: Muroidea – myszowe
- rodzina: Platacanthomyidae – kolcosznicowate
- rodzina: Spalacidae – ślepcowate
- rodzina: Calomyscidae – myszochomikowate
- rodzina: Nosomyidae – malgaszomyszowate
- rodzina: Cricetidae – chomikowate
- rodzina: Muridae – myszowate
- infrarząd: Castorimorphi – bobrokształtne
- podrząd: Hystricomorpha – jeżozwierzowce
- infrarząd: Ctenodactylomorphi – gundiokształtne
- rodzina: Ctenodactylidae – gundiowate
- rodzina: Diatomyidae – gundioszczurowate
- infrarząd: Hystricognathi – jeżozwierzokształtne
- rodzina: Hystricidae – jeżozwierzowate
- rodzina: Thryonomyidae – szczecińcowate
- rodzina: Petromuridae – skałoszczurowate
- rodzina: Heterocephalidae – golce
- rodzina: Bathyergidae – kretoszczurowate
- nadrodzina: Erethizontoidea
- rodzina: Erethizontidae – ursonowate
- nadrodzina: Cavioidea
- rodzina: Cuniculidae – pakowate
- rodzina: Caviidae – kawiowate
- rodzina: Dasyproctidae – agutiowate
- nadrodzina: Chinchilloidea
- rodzina: Chinchillidae – szynszylowate
- rodzina: Dinomyidae – pakaranowate
- nadrodzina: Octodontoidea
- rodzina: Abrocomidae – szynszyloszczurowate
- rodzina: Ctenomyidae – tukotukowate
- rodzina: Octodontidae – koszatniczkowate
- rodzina Echimyidae – kolczakowate
- infrarząd: Ctenodactylomorphi – gundiokształtne
- podrząd: Sciuromorpha – wiewiórkokształtne
- rodzina: Aplodontiidae – sewelowate
- rodzina: Sciuridae – wiewiórkowate
- rodzina: Gliridae – popielicowate
Taksony wymarłe należące do rzędu Rodentia o niepewnej pozycji systematycznej i nie zgrupowane w żadnyn z powyżśzych podrzędów:
Rodziny:
- Archetypomyidae Meng, Li, Ni, Wang Yuanqing & Beard, 2007
- Cylindrodontidae G.S. Miller & Gidley, 1918
- Ivanantoniidae Shevyreva, 1989
- Laredomyidae J.A. Wilson & Westgate, 1991
- Protoptychidae Schlosser, 1911
- Remyidae Hartenberger, 1973
- Sciuravidae G.S. Miller & Gidley, 1918
- Zelomyidae Dawson, Huang, Li & Wang Banyue, 2003
- Zetamyidae Ferrusquía-Villafranca, Flynn, Ruiz-González, Torres-Hernández & Martínez-Hernández, 2018
Rodzaje:
- Corbarimys Marandat, 1989
- Dolosimus Dawson, Li & Qi, 2010
Gryzonie i człowiek
Ochrona
Gryzonie nie zaliczają się do najbardziej zagrożonych grup ssaków. Jednak 168 gatunków należących do 126 rodzajów wymaga działań ochronnych w obliczu ograniczonego uznania tego przez opinię publiczną. 76% rodzajów gryzoni to taksony monotypowe, więc zmienność filogenetyczna może ubożeć przy względnie niewielkiej liczbie wyginięć. Przy braku szczegółowej wiedzy na temat zagrożonych gatunków i dokładnej taksonomii ochrona musi bazować na wyższych taksonach, raczej w randze rodzin niż poszczególnych gatunków, i geograficznych gorących punktach. Kilka gatunków ryżniaka (Oryzomys) wyginęło od XIX wieku, prawdopodobnie na skutek utraty siedlisk i introdukcji obcych gatunków. Pochodzący z Kolumbii igłozwierz kolumbijski odnotowany został jedynie w dwóch lokalizacjach w latach dwudziestych XX wieku, natomiast szybowniczek rudogrzbiety znany jest jedyne z lokalizacji typowej na wybrzeżu karaibskim. Gatunkom tym przysługuje status narażonych (VU – vulnerable). Komisja IUCN podnosi, że można bezpiecznie stwierdzić, iż wiele południowoamerykańskich gryzoni jest poważnie zagrożonych. Jako główne powody takiego stanu podaje niszczenie środowiska naturalnego i intensywne polowania.
Z drugiej strony mówi się o trzech nowych kosmopolitycznych komensalnych gryzoniach-szkodnikach. Chodzi o szczura śniadego, szczura wędrownego i mysz domową. Rozprzestrzeniły się one razem z człowiekiem, po części na statkach handlowych za czasów wielkich odkryć geograficznych. Wraz z czwartym gatunkiem działającym w obrębie Pacyfiku, szczurem polinezyjskim, poważnie zniszczyły bioty wysp na świecie. Przykładowo kiedy szczur śniady trafił na wyspę Lord Howe w 1918, ponad 40% lądowych gatunków ptaków zamieszkujących wyspę, w tym podgatunku wachlarzówki posępnej Rhipidura fuliginosa cervina, wyginęło w przeciągu następnych 10 lat. Podobna katastrofa zdarzyła się na Midway w 1943 i Big South Cape Island w 1962. Projekty ochronne mogą przy starannym planowaniu całkowicie wyplenić te szkodniki z wysp, wykorzystując w tym celu antykoagulanty, rodentycydy takie jak brodifacoum. Udało się to na wyspie Lundy w Wielkiej Brytanii, gdzie eradykacja około 40 000 szczurów wędrownych dała szansę burzykowi północnemu i maskonurowi zwyczajnemu na odbudowę populacji, znajdujących się na krawędzi zagłady.
Wykorzystanie

Ludzkość od dawna używała zwierzęcych skór do produkcji ubrań, jako że skóra jest trwała, a futro zapewnia dodatkową izolację. Rdzenni mieszkańcy Ameryki Północnej używali często skór bobrów, garbując je i zszywając razem, czyniąc z nich szaty. Europejczycy docenili ich jakość i tak rozwinął się północnoamerykański rynek futra, stając się najważniejszym dla wczesnych sprzedawców. W Europie miękki podszerstek znany jako wełna z bobra uznano za idealny do filcowania, służył do produkcji nakryć głowy i ubrań. Później przerzucono się na tańszą nutrię, która również służyła do produkcji filcu. Rozlegle hodowano ją na farmach Ameryki i Europy. Jednak moda zmieniła się, dostępne stały się nowe materiały i ta dziedzina przemysłu skurczyła się. Szynszyla ma miękki i jedwabisty podszerstek. Zapotrzebowanie nań urosło do takich rozmiarów, że zwierzę prawie wytępiono na wolności, zanim głównym źródłem skór stały się farmy. Kolce i sierść ursonowatych służą tradycyjnej dekoracji ubrań. Przykładowo ich sierść służy tworzeniu przez aborygenów amerykańskich pióropuszy. Główne kolce można ufarbować i następnie wkomponować nitki, by upiększyć wyroby skórzane, jak pochwy noży czy torby. Dakotańskie kobiety zbierają kolce dla wyroby specjalnych tekstyliów. Rzucają koc na ursona i zbierają pozostawione przezeń w materiale kolce.
Przynajmniej 89 gatunków gryzoni, w większości przedstawicieli Hystricomorpha, jak kawie domowe (świnki morskie), aguti i kapibary, są jadane przez ludzi. W 1985 istniały przynajmniej 42 różne ludzkie społeczności, których członkowie jedli szczury. Kawie domowe po raz pierwszy posłużyły człowiekowi za pokarm około 2500 p.n.e. i do 1500 p.n.e. stały się głównym źródłem mięsa w Imperium Inków. Rzymianie hodowali popielice w glirariach lub na dużych ogrodzonych terenach na dworze. Tuczono je orzechami włoskimi, kasztanami i żołędziami. Popielice łapano także w stanie dzikim jesienią, kiedy były najtłustsze, by piec je po zanurzeniu w miodzie lub nadzianiu wieprzowiną, orzeszkami piniowymi czy innymi przyprawami. W Amazonii, gdzie dokucza niedobór dużych ssaków, pakowate i aguti stanowiły około 40% zwierzyny spożywanej przez Indian, ale na obszarach leśnych obfitujących w wielkie ssaki wymienione gryzonie stanowiły tylko 3% zdobyczy.
Kawie domowe wykorzystuje kuchnia Cusco (Peru), w daniu nazywanym cuy al horno (pieczona kawia domowa, czyli świnka morska). Tradycyjny piec andyjski zwany qoncha lub fogón robiony jest z błota i gliny wzmocnionych słomą i włosiem zwierząt takich jak kawia domowa. W Peru istnieje stała populacja domowych kawii domowych 20 milionów sztuk, pozwalająca na pozyskanie 64 milionów tusz przeznaczonych do konsumpcji. Zwierzę to jest świetnym źródłem pożywienia, zawiera 19% białka. W USA zjada się głównie wiewiórki, ale też piżmaki, ursonowate i świstaki amerykańskie. Nawahowie spożywają nieświszczuki płowe pieczone w błocie, podczas gdy Pajutowie jedzą gofferowate, wiewiórki i szczury.
Takie gryzonie, jak kawie domowe, myszy, szczury, chomiki, suwaki, szynszyle, koszatniczka i pręgowiec, stanowią dogodne zwierzęta domowe, aczkolwiek każdy gatunek jest inny. Większość normalnie trzyma się w klatkach odpowiednich rozmiarów – wymagania przestrzenne i społeczne są zróżnicowane. Trzymane w rękach od maleńkości, zazwyczaj zachowują się łagodnie i nie gryzą. Kawia domowa żyje dłużej i potrzebuje też większej klatki. Szczur również wymaga sporej przestrzeni. Może się całkiem oswoić, uczyć sztuczek i wydaje się zadowolony z towarzystwa człowieka. Mysz żyje krótko i potrzebuje bardzo niewiele miejsca. Chomik wiedzie samotny, ale nocny tryb życia. Wykazuje interesujące zachowania, jeśli jednak nie bierze się go w rękę regularnie, broni się. Suwaki zazwyczaj nie wykazują agresji, rzadko jednak gryzą. Są też zwierzętami społecznymi i lubią towarzystwo człowieka i innych osobników.

Gryzonie są szeroko wykorzystywane jako organizmy modelowe w badaniach na zwierzętach. Albinotyczny mutant szczura laboratoryjnego po raz pierwszy użyty został w badaniach w 1828. Później stał się pierwszym zwierzęciem udomowionym dla celów ściśle naukowych. Współcześnie laboratoria najczęściej z gryzoni wykorzystują mysz domową. W 1979 liczbę wykorzystywanych osobników oszacowano na 50 milionów rocznie. Wybiera się je z uwagi na małe rozmiary, płodność, krótką ciążę, łatwość chwytania ręką oraz wrażliwość na wiele stanów chorobowych nękających również ludzi. Wykorzystuje się je w genetyce, biologii rozwoju, cytologii, onkologii i immunologii. Popularne w laboratoriach do końca XX wieku były też kawie domowe (świnki morskie). W latach sześćdziesiątych w USA używano rocznie 2,5 miliona osobników, ale do lat dziewięćdziesiątych liczba ta spadła do około 375 tysięcy. W 2007 stanowiły one około 2% zwierząt laboratoryjnych. Odegrały główną rolę w odkryciu roli mikroorganizmów w patogenezie chorób pod koniec XIX stuleciu, dzięki eksperymentom Pasteura, Rouxa i Kocha. Kilka razy wyniesiono je również w przestrzeń kosmiczną. ZSRR użyło do tego biosatelity Sputnika 9 w dniu 9 marca 1961. Akcja zakończyła się sukcesem. Golec to jedyny znany ssak poikilotermiczny. Wykorzystuje się go w badaniach termoregulacji. Wyróżnia się też brakiem produkcji jednego z neuroprzekaźników, substancji P. Fakt ten wykorzystuje się w badaniach bólu.
Rodentia cechują się czułym zmysłem węchu. Używano ich więc do wykrywania zapachów czy poszukiwanych substancji. Wielkoszczur potrafi wykryć wywołujące gruźlicę prątki z czułością do 86,6% i swoistością ponad 93%. Osobniki tego samego gatunku uczono wykrywać miny lądowe. Szczury badano pod kątem użycia w niebezpiecznych sytuacjach, jak strefy katastrof. Zwierzęta te mogą nauczyć się odpowiadać na komendy, które można wydawać z oddali. Nakłoniono je nawet do odwiedzenia oświetlonych jasnych miejsc, których szczury zazwyczaj unikają.
Szkodniki i przenosiciele chorób

Niektóre gatunki gryzoni uznaje się za poważne szkodniki w rolnictwie, zjadające duże ilości jedzenia przeznaczonego dla ludzi. Przykładowo w 2003 ilość ryżu straconego przez myszy i szczury w Azji wedle szacunków mogłaby nakarmić 200 milionów ludzi. Większość szkód na całym świecie wyrządza stosunkowo mała grupa gatunków, głównie myszy i szczurów. W Indonezji i Tanzanii gryzonie zmniejszają plony o około 15%, ale w niektórych wypadkach w Ameryce Południowej nawet o 90%. W Afryce gryzonie, w tym przedstawiciele rodzajów myszorówka i Kusu, niszczą zboża, orzechy ziemne, warzywa i kakao. W Azji myszy, szczury i gatunki takie jak chinornik stepowy, suwak mongolski i Eospalax baileyi niszczą zbiory ryżu, sorgo, bulw, warzyw i orzechów. W Europie oprócz szczurów i myszy, myszarka, nornik, a podczas okazjonalnych wzrostów liczebności także karczownik ziemnowodny, powodują szkody w sadach, warzywach, na pastwiskach, jak i w zbożach. W Ameryce Południowej szkody powoduje szerszy wachlarz gatunków, choćby z rodzajów płetwoszczurek, trawniak, wieczorniczka, ryżaczek, liściouch, bawełniak i szuwarniczka. Niszczą one wiele plonów, w tym także trzcinę cukrową, owoce, warzywa i bulwy.
Gryzonie stanowią także ważne wektory chorób. Szczur śniady, wraz z przenoszonym przezeń pchłami Nosopsyllus fasciatus odgrywa pierwszorzędową rolę w roznoszeniu bakterii wywołujących dżumę (Yersinia pestis). Przenosi także mikroorganizmy wywołujące tyfus plamisty, leptospirozy, toksoplazmozę i włośnicę. Wiele gryzoni roznosi hantawirusy, jak wirus Puumala, wirus Dobrava czy wirus Saaremaa, mogące wywołać infekcję u człowieka. Gryzonie biorą także udział w transmisji takich chorób, jak: babeszjoza, leiszmanioza skórna, anaplazmoza, borelioza, omska gorączka krwotoczna, choroby wywoływane przez wirus Powassan i wirus Zachodniego Nilu, riketsjozy, dur powrotny, gorączka plamista Gór Skalistych.
Społeczności ludzkie często podejmowały działania w celu kontroli gryzoni, szkodników i zwierząt stwarzających zagrożenie dla zdrowia publicznego. Ludzie tradycyjnie zastawiali trutki i pułapki. Metody nie zawsze były bezpieczne czy efektywne. Bardziej współcześnie integrowana ochrona stara się poprawić kontrolę populacji gryzoni dzięki połączeniu metod ustalenia wielkości i rozmieszczenia populacji szkodników, utworzenia limitów tolerancji (poziomu aktywności szkodnika, przy której rozpoczyna się interwencję), samej interwencji oraz ewaluacji efektywności bazującej na powtarzalnych metodach. Interwencje mogą obejmować edukację, wprowadzanie i przystosowywanie prawa i innych regulacji, modyfikację siedlisk, zmianę praktyk rolniczych, kontrolę biologiczną przy użyciu patogenów lub drapieżników, jak też rodentycydów i pułapek. Wykorzystanie patogenów takich, jak Salmonella, ma wady. Mogą one zainfekować człowieka bądź zwierzęta przezeń udomowione. Gryzonie natomiast mogą nabyć oporność. Wykorzystuje się też drapieżniki: fretki domowe, mangustowate i waranowate. Efekty nie są jednak satysfakcjonujące. Koty udomowione bądź dzikie potrafią efektywnie kontrolować populacje gryzoni, o ile nie są one zbyt liczebne. W Wielkiej Brytanii szczególnie dwa gatunki, mysz domowa i szczur wędrowny, są aktywnie kontrolowane w celach ograniczenia zniszczeń wzrastających plonów, kontaminacji zapasów z tychże, a także zniszczeń w infrastrukturze, takimi sposobami, by zachować zgodność z prawem.
Uwagi
Bibliografia
- J.K.W. Illiger: Prodromus systematis mammalium et avium: additis terminis zoographicis utriusque classis, eorumque versione germanica. Berolini: Sumptibus C. Salfeld, 1811, s. 1–301. (łac.).
- S.O. Landry. A proposal for a new classification and nomenclature for the Glires (Lagomorpha and Rodentia). „Mitteilungen aus dem Museum für Naturkunde in Berlin”. Zoologische Reihe. 75 (2), s. 283–316, 1999. DOI: 10.1002/mmnz.19990750209. (ang.).