

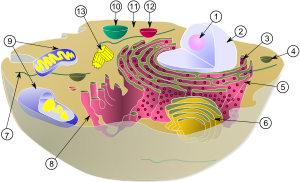
1) jąderko
2) jądro komórkowe
3) rybosom
4) pęcherzyk
5) szorstkie retikulum endoplazmatyczne
6) aparat Golgiego
7) mikrotubule
8) gładkie retikulum endoplazmatyczne
9) mitochondrium
10) wakuola
11) cytozol
12) lizosom
13) centriola
Mitochondrium (w liczbie mnogiej mitochondria) – otoczone dwiema błonami organellum, obecne w większości komórek eukariotycznych. Organella te mają różną wielkość, przeważnie od 2 do 8 μm, mogą też szybko zmieniać swój kształt i rozmiary. Są one miejscem, w którym w wyniku procesu oddychania komórkowego powstaje większość adenozynotrifosforanu (ATP) komórki, będącego jej źródłem energii. Oprócz tego mitochondria są zaangażowane w wiele innych procesów, takich jak sygnalizacja komórkowa, specjalizacja, wzrost i śmierć komórki, czy też kontrola cyklu komórkowego. Nazwa pochodzi od greckiego μίτος (mítos) – "nić" oraz χονδρίον (chondríon) – "ziarenko".
Kilka cech mitochondriów czyni je unikatowymi pośród organelli. Ich liczba w pojedynczej komórce jest bardzo różna w zależności od organizmu i typu komórki. Przeciętna komórka eukariotyczna zawiera od kilkuset do kilku tysięcy mitochondriów. Wiele komórek jednakże ma tylko jedno mitochondrium, u kilku zaś, na przykład u ameby Chaos chaos L., stwierdzono po kilka tysięcy mitochondriów. Nowe mitochondria powstają zwykle poprzez wzrost i podział już istniejących (większość białek mitochondrium jest kodowana przez DNA jądrowe). Organellum to jest złożone z kilku przedziałów, mających specyficzne funkcje. Te przedziały to błona zewnętrzna, przestrzeń międzybłonowa, błona wewnętrzna, grzebienie oraz macierz mitochondrialna. Białka mitochondrialne mogą być różne, w zależności od komórki i gatunku. W mitochondriach ludzkiego serca zostało zidentyfikowanych 615 różnych rodzajów białek, podczas gdy u szczurów liczba ta wynosi 940.
Pomimo tego, że większość genomu komórki znajduje się w jądrze komórkowym, mitochondria, jako jedyne organella poza plastydami, mają własny genom. Genom mitochondrialny jest nieduży – koduje tylko od kilkunastu do kilkudziesięciu białek z kilkuset białek niezbędnych do funkcjonowania mitochondrium. Co więcej, wykazuje on podobieństwo do genomu bakterii.
Budowa

1) membrana wewnętrzna
2) membrana zewnętrzna
3) grzebień mitochondrialny
4) matrix mitochondrialna

Mitochondrium składa się z dwóch błon; zewnętrznej i wewnętrznej, zbudowanych z dwuwarstwy lipidowej oraz rozmieszczonych w niej białek. Są one podobne w budowie do zwykłej błony komórkowej, jednak obydwie błony mają odmienne właściwości. Z powodu takiej budowy, w mitochondrium można wyróżnić pięć odrębnych przedziałów. Są to: błona mitochondrialna zewnętrzna, przestrzeń międzybłonowa (pomiędzy błoną zewnętrzną a wewnętrzną), błona mitochondrialna wewnętrzna, grzebienie mitochondrialne (tworzone przez fałdy błony wewnętrznej) oraz macierz mitochondrialną (wewnętrzna przestrzeń mitochondrium).
Błona zewnętrzna
Błona zewnętrzna mitochondrium otacza organellum, oddzielając je od środowiska zewnętrznego. Ma ona współczynnik białek błonowych do lipidów podobny jak u większości błon komórkowych eukariontów (około 1:1 wagowo). Zawiera duże ilości białek integralnych zwanych porynami. Poryny są w istocie dużymi (około 2-3 nm średnicy) kanałami, przez które mogą się swobodnie przedostawać wszystkie białka o masie mniejszej niż 5000 daltonów. Białka o większych cząsteczkach mogą pokonać błonę zewnętrzną tylko za pomocą transportu aktywnego. Ich N-koniec wiąże się wtedy z podjednostką białka zwanego translokazą błony zewnętrznej, która przenosi je na drugą stronę błony. Małe cząsteczki, na przykład woda czy dwutlenek węgla mogą swobodnie dyfundować przez tę błonę. Przerwanie błony zewnętrznej skutkuje uwolnieniem do cytozolu białek znajdujących się w przestrzeni międzybłonowej, co prowadzi do śmierci komórki. Błona zewnętrzna może łączyć się z retikulum endoplazmatycznym, tworząc strukturę zwaną MAM (mitochondria-associated ER-membrane (ang.)). Jest to ważne przy wydalaniu przez retikulum jonów wapnia oraz pełni rolę przy transporcie lipidów z retikulum do mitochondrium.
Wśród białek budujących błonę zewnętrzną mitochondrium występują enzymy odpowiadające za bardzo rozmaite reakcje, jak np. wydłużanie łańcuchów kwasów tłuszczowych, utlenianie adrenaliny i rozkład tryptofanu. Enzymem markerowym (markerem) błony zewnętrznej jest oksydaza monoaminowa (MAO).
Przestrzeń międzybłonowa
Przestrzeń międzybłonowa to przestrzeń pomiędzy zewnętrzną a wewnętrzną błoną mitochondrium. Ponieważ błona zewnętrzna jest przepuszczalna dla małych cząsteczek, stężenie substancji takich jak jony lub cukry w przestrzeni międzybłonowej jest takie samo jak w cytozolu. Duże białka jednak, muszą mieć specjalne sekwencje sygnałowe aby zostać przetransportowane przez błonę zewnętrzną, ich skład w przestrzeni międzybłonowej jest odmienny od tego, występującego w cytozolu. Jednym z białek charakterystycznych dla tej przestrzeni jest cytochrom c, zaś jej enzymem markerowym (markerem) jest kinaza adenilanowa.
Błona wewnętrzna
Błona wewnętrzna mitochondrium, to błona znajdująca się w środku organellum. To właśnie w niej zachodzą reakcje chemiczne, przekształcające energię cząsteczek substancji pokarmowych w energię wiązań ATP. Ogólnie biorąc, w błonie tej występuje pięć typów białek o różnych funkcjach: białka biorące udział w reakcjach redoks fosforylacji oksydacyjnej, syntaza ATP wytwarzająca ATP w macierzy mitochondrialnej, specjalne białka kontrolujące przechodzenie metabolitów do oraz z macierzy, białka zajmujące się importem innych białek oraz te, które zajmują się syntezą oraz rozkładem innych białek.
W jej skład wchodzi ponad 150 polipeptydów, ma także wysoki współczynnik białek do lipidów (ponad 3:1 wagowo, oznacza to, że 1 białko przypada na 15 fosfolipidów). Są to białka łańcucha oddechowego, syntaza ATP wytwarzająca ATP w macierzy mitochondrialnej oraz białka transportujące metabolity do wnętrza macierzy i na zewnątrz. Stanowią one około 20% wszystkich białek mitochondrium. Błona wewnętrzna, w przeciwieństwie do błony zewnętrznej nie zawiera poryn i jest nieprzepuszczalna dla wszystkich cząsteczek. Posiada ona jednakże w swojej strukturze nietypowy fosfolipid, kardiolipinę. Związek ten, odkryty po raz pierwszy w krowim sercu w 1942, jest charakterystyczny dla błon bakterii i mitochondrium. Kardiolipina zawiera w swojej strukturze cztery nasycone kwasy tłuszczowe zamiast dwóch, co może powodować, że nasycona nią błona będzie trudniejsza do spenetrowania. Transport jonów i innych cząsteczek dostających się oraz wychodzących z macierzy mitochondrialnej wymaga specjalnych transporterów błonowych. Białka przenoszone są przez kompleks translokazy błony wewnętrznej mitochondrium lub przez Oxa1. Umożliwia to wytworzenie gradientu protonowego niezbędnego do działania łańcucha oddechowego.
Grzebienie
Powierzchnia błony wewnętrznej mitochondrium, na przykład w mitochondriach wątroby, jest pięciokrotnie większa od powierzchni zewnętrznej błony mitochondrialnej. Z tego powodu wpukla się ona do środka, tworząc charakterystyczne wpuklenia, tak zwane grzebienie mitochondrialne. Wpuklenia zwiększają powierzchnię błony wewnętrznej, powiększając znacznie obszar, na którym zachodzi produkcja ATP. Powierzchnia ta nie jest stała i mitochondria komórek które mają większe zapotrzebowanie na ATP, takie jak komórki mięśni, tworzą grzebienie o większej powierzchni. Grzebienie usiane są małymi, okrągłymi ciałami, zwanymi oksysomami. Nie są to wpuklenia przypadkowe, a raczej wytwory błony zewnętrznej, których zadaniem jest kontrolowanie chemiosmozy. W grzebieniach zakotwiczone są enzymy łańcucha oddechowego.
Macierz mitochondrialna
Macierz mitochondrialna, inaczej matriks, to przestrzeń wewnątrz mitochondrium, ograniczona błoną wewnętrzną. Wypełnia ją rodzaj żelu – wodny roztwór białek i metabolitów zużywanych na potrzeby mitochondrium. Macierz zawiera około 2/3 wszystkich białek w mitochondrium. W ich skład wchodzą takie białka jak enzymy β-oksydacji kwasów tłuszczowych, cyklu Krebsa, syntezy steroidów i inne. Enzymem markerowym (markerem) matrix mitochondrialnej jest syntetaza cytrynianowa. Macierz zawiera również materiał genetyczny w postaci kilku kopii mitochondrialnego DNA (mtDNA), rybosomy mitochondrialne i tRNA mitochondrialny.
Organizacja i występowanie
Mitochondria występują u prawie wszystkich eukariotów. Ich kształt i liczba zmieniają się w zależności od organizmu, typu komórki oraz jej zapotrzebowania na energię. Pojedyncza komórka zawiera od kilku sztuk do kilku tysięcy mitochondriów, przeciętnie kilkaset. U organizmów jednokomórkowych często obecne jest tylko jedno mitochondrium, z drugiej strony w komórkach wątroby ssaków można znaleźć około 1500 mitochondriów, stanowiących do 1/5 objętości komórki. W komórkach roślinnych, niezróżnicowanych komórkach zwierzęcych, komórkach regenerujących, limfocytach oraz w komórkach naskórka występuje po kilkaset tych organelli. Szczególnie dużo, ok. 1-2 tysięcy mitochondriów, występuje w komórkach wątrobowych, komórkach gruczołów żołądkowych, kanalików nerkowych krętych i komórkach kory nadnerczy czy komórkach tkanki mięśniowej typu sercowego.
Mitochondria nie występują w końcowym okresie różnicowania się erytrocytów (u ssaków) czy komórek soczewki.
Mitochondria przyjmują zwykle okrągły lub owalny kształt, istnieją jednak również mitochondria o kształcie niciowatym i rozgałęzionym. Takie formy występują na przykład u jednokomórkowych wiciowców, glonów i drożdży. U zwierząt obecność form niciowatych stwierdzono w komórkach trzustki, wstawce plemnika, a także w komórkach wątroby. Wraz z cytoszkieletem tworzą one rozgałęzioną, trójwymiarową sieć złączonych ze sobą mitochondriów. Takie połączenie może wpływać na przepuszczalność błony zewnętrznej mitochondrium dla ADP.
Lokalizacja mitochondriów w komórce nie jest stała. W wyniku ruchów cytoplazmy lub dzięki związaniu się z elementami cytoszkieletu, mitochondria mają zdolność do przemieszczania się w kierunku miejsca o zwiększonym zapotrzebowaniu na energię. Mogą one lokować się na przykład między fibrylami komórek mięśniowych, w aparacie kurczliwym mięśnia sercowego, u podstawy komórek nabłonka gruczołowego, w zakończeniach włókien nerwowych – synapsach, wzdłuż włókien wrzeciona cytokinetycznego, u podstawy witki w plemniku lub w pobliżu substratów oddechowych takich jak krople tłuszczu.
Funkcja
Najważniejszymi rolami mitochondriów są wytwarzanie ATP poprzez oddychanie komórkowe oraz regulacja metabolizmu komórki. Główny szereg reakcji biochemicznych związany z produkcją ATP, u eukariontów zachodzący wyłącznie w mitochondriach, nazywany jest cyklem kwasu cytrynowego lub cyklem Krebsa. Są to najważniejsze reakcje mające miejsce w mitochondriach, jednak poza nimi mitochondria pełnią także inne funkcje.
Przemiany energetyczne
Najważniejszą rolą mitochondrium jest wytwarzanie ATP, znajdujące odzwierciedlenie w ilości białek błony wewnętrznej mitochondrium, które je przeprowadzają. Zachodzi ono dzięki utlenianiu głównych produktów rozkładu glukozy – pirogronianu i NADH, wytwarzanych w cytozolu. Ten proces oddychania komórkowego, nazywanego także oddychaniem tlenowym, zależny jest od obecności tlenu. Kiedy ilość tlenu dostarczanego mitochondriom jest ograniczona, produkty glikolizy przetwarzane są w ramach oddychania beztlenowego, procesu który nie zachodzi w mitochondriach. Jest to jednakże proces niekorzystny z energetycznego punktu widzenia, ponieważ podczas oddychania tlenowego uzyskiwane jest około 13 razy więcej energii niż podczas oddychania beztlenowego. Ostatnio zostało dowiedzione, że mitochondria roślinne mogą wytworzyć pewną ilość ATP nawet bez tlenu, stosując jako substrat azotyny.
Cykl kwasu cytrynowego
Cząsteczki pirogronianu, które powstają w wyniku glikolizy, są aktywnie transportowane poprzez błonę wewnętrzną mitochondrium do macierzy mitochondrialnej, gdzie zostają utlenione i połączone z koenzymem A tak, by powstał CO2, acetylokoenzym A oraz NADH. Acetylokoenzym A jest pierwszym substratem cyklu kwasu cytrynowego. Enzymy tego cyklu zlokalizowane są głównie w macierzy mitochondrialnej, z wyjątkiem dehydrogenazy bursztynianowej, która umocowana jest w błonie wewnętrznej mitochondrium, gdzie wchodzi w skład kompleksu II. Cykl kwasu cytrynowego utlenia acetylo-CoA do dwutlenku węgla, a także powoduje powstanie zredukowanych kofaktorów: trzech cząsteczek NADH i dwóch cząsteczek FADH2, stanowiących źródło elektronów dla łańcucha oddechowego, oraz cząsteczki GTP, która od razu przemieniana jest w ATP.
Łańcuch oddechowy
Potencjał redoks pochodzący od NADH i FADH2, służący do uzyskania energii podczas utleniania, jest przekazywany tlenowi stopniowo, poprzez łańcuch oddechowy. Te bogate w energię cząsteczki powstają w macierzy mitochondrialnej podczas cyklu kwasu cytrynowego oraz w cytoplazmie komórki podczas glikolizy. Reduktory z cytoplazmy mogą być importowane do mitochondrium przez antyportowe czółenko jabłczanowo-asparaginowe lub przy użyciu czółenka glicerofosforanowego. Transport ten przeprowadzają kompleksy białkowe umieszczone w błonie wewnętrznej mitochondrium, takie jak dehydrogenaza NADH, cytochrom bc1 i oksydaza cytochromu c. Uzyskana energia używana jest do pompowania protonów (H+) do przestrzeni międzybłonowej. Proces ten jest wydajny, jednak pewna część elektronów redukuje tlen przedwcześnie, powodując powstanie reaktywnych form tlenu, takich jak ponadtlenki. Stanowi to przyczynę stresu oksydacyjnego oraz może przyczynić się do podupadania funkcji mitochondriów związanego z procesem starzenia się.
Gdy stężenie protonów w przestrzeni międzybłonowej wzrasta, po przeciwnych stronach błony wewnętrznej wytwarza się silny gradient elektrochemiczny. Jest on powodowany parciem protonów do powrotu do macierzy mitochondrialnej. Jedyną drogą powrotu jest kompleks białkowy syntazy ATP. Energia potencjalna przechodzących przez niego protonów używana jest do syntezy ATP z ADP oraz anionu fosforanowego (Pi). Proces ten nazywany jest chemiosmozą.
Produkcja ciepła
W pewnych warunkach protony mogą przedostawać się do macierzy mitochondrialnej, nie wytwarzając ATP. Proces ten określany jest mianem wyciekaniem protonów bądź mitochondrialne rozprzęgnięcie, zachodzi zaś dzięki dyfuzji wspomaganej protonów do macierzy. Powoduje to rozproszenie energii potencjalnej gradientu elektrochemicznego protonów w postaci ciepła. Proces przeprowadzany jest dzięki działaniu kanału protonowego zwanego termogeniną lub UCP1. Termogenina jest białkiem o masie 33 kDa, odkrytym w 1973 roku. Znajdywana jest głównie w gruczołach snu zimowego, zwanych także tłuszczem brunatnym, odpowiedzialnych za produkcję ciepła niezwiązaną z drżeniem. Tkanka ta obecna jest u ssaków, zwłaszcza młodych, bądź u gatunków, które odbywają sen zimowy. U ludzi ilość tłuszczu brunatnego największa jest tuż po urodzeniu, potem zaś maleje z wiekiem.
Magazynowanie jonów wapnia
Ilość wolnego wapnia w komórce może regulować szereg reakcji oraz jest bardzo ważna dla przewodnictwa sygnałów w komórce. Mitochondria mogą przejściowo magazynować wapń, co stanowi część procesów odpowiedzialnych za zachowanie równowagi wapniowej w komórce. Ich zdolność do szybkiego przyjmowania wapnia w celu późniejszego uwolnienia czyni je dobrymi buforami równowagi wapniowej. Główną rolę w magazynowaniu wapnia pełni retikulum endoplazmatyczne, a między nim a mitochondrium dochodzi do znaczących interakcji pod kątem gospodarki wapniowej. Wapń przechodzi swobodnie przez błonę zewnętrzną mitochondrium do przestrzeni międzybłonowej, skąd transportowany jest do macierzy mitochondrialnej za pomocą uniportu wapniowego w błonie wewnętrznej. Transport ten napędzany jest przez potencjał błonowy mitochondrium. Uwolnienie wapnia z powrotem do wnętrza komórki może zajść dzięki białku wymiany sodowo wapniowej bądź dzięki ścieżce wapniowo indukowanego uwalniania wapnia. Może to spowodować nagłe lub falowe zmiany potencjału błonowego, co może wywołać reakcję w postaci wypuszczenia neurotransmiterów bądź hormonów.
Inne funkcje
Mitochondria pełnią role także w innych procesach metabolicznych, takich jak:
- regulowanie potencjału błonowego
- apoptoza – programowana śmierć komórki
- regulacja stanu redoks komórki
- synteza hemu
- synteza sterydów
- cykl mocznikowy – w mitochondriach wątroby.
Pochodzenie


Obecnie najszerzej przyjmowany pogląd głosi, że mitochondria są potomkami pewnych organizmów endosymbiotycznych, podobnych do dzisiejszych bakterii, które w jakiś sposób przetrwały endocytozę do innej komórki i zostały włączone w skład pierwotnych organizmów eukariotycznych. Teoria endosymbiozy po raz pierwszy została zaproponowana przez Konstantina Mereszkowskiego na początku XX wieku. Pogląd ten został po pewnych zmianach spopularyzowany w latach 70. XX wieku przez Lynn Margulis. Możliwość przeprowadzania tlenowego oddychania komórkowego w mitochondriach, dzięki któremu możliwe byłoby uzyskanie większej ilości energii z tej samej ilości pokarmu niż u innych organizmów, stanowiła dużą przewagę ewolucyjną i przyczyniła się do sukcesu ewolucyjnego organizmów mających mitochondria. Zwiększyło to także liczbę środowisk, w których takie organizmy mogły się rozwijać. Ocenia się, że do tej endosymbiozy doszło około 2 do 1,7 miliarda lat temu.
Mitochondria mają wiele cech, które występują także u prokariotów. Po pierwsze, mitochondrium zawiera DNA, który zorganizowany jest w postaci kilku kopii pojedynczego, kołowego nukleoidu, podobnie jak to ma miejsce u prokariotów. Co więcej, kod genetyczny mitochondriów jest kodowany w podobny sposób, jaki ma miejsce u proteobakterii. Także rybosomy kodowane przez mitochondrialny DNA przypominają kształtem i wielkością te, które można spotkać u prokariotów. Mają one wielkość 70S, taką samą jak rybosomy bakteryjne, nie zaś 80S, jaką mają rybosomy występujące w cytozolu komórki eukariotycznej. Świadczy to o tym, że przodkiem mitochondriów był organizm należący do proteobakterii. Niektóre badania sugerują, że należał on do riketsji, jednak sprawa pochodzenia mitochondriów i ich pokrewieństwa do proteobakterii pozostaje kontrowersyjna.
Według innej teorii mitochondria mogły powstać w tym samym czasie, w którym doszło do wytworzenia błony jądrowej, i mogą być efektem tego samego procesu, który doprowadził do wyodrębnienia jądra komórkowego.
Istnieje kilka grup jednokomórkowych organizmów eukariotycznych niemających mitochondriów, na przykład mikrosporydia, Metamonada, Archamoebae. U wielu z nich jednak zachowały się szczątkowe organella będące ewolucyjną pozostałością mitochondriów – mitosomy. Część anaerobowych eukariontów wykorzystuje organella homologiczne do mitochondriów do produkcji związków siarkowo-żelazowych biorących udział w gospodarce białkowej. Według stanu wiedzy z 2016 r. jedynym eukariontem, który nie ma ani mitochondriów lub ich homologów, ani nawet genów pochodzenia mitochondrialnego jest przedstawiciel rodzaju Monocercomonoides.
Dziedziczenie
U ssaków mitochondria płodu pochodzą wyłącznie z komórki jajowej (plemnik, tworząc przedjądrze męskie, pozostawia wszystkie swoje organella poza komórką jajową), u wielu innych organizmów (np. owady) plemnik wnika jednak do komórki jajowej razem z własnymi mitochondriami. W związku z tym mitochondria dziedziczymy wyłącznie w linii matczynej, „po kądzieli”, a geny mitochondrialne nie ulegają rearanżacji przez rekombinację. Z tego powodu geny mitochondrialne porównywano dla ustalenia kiedy żyła kobieta, od której pochodzą wszystkie aktualne mitochondria, nazwana niefortunnie przez prasę Ewą mitochondrialną, co mylnie sugeruje jedynego przodka wszystkich ludzi. Wyniki wskazują na ok. 200 tysięcy lat i Afrykę, z dużym marginesem błędu (kilkadziesiąt tysięcy lat). Wyniki te, podobnie jak wiele innych badań genetycznych, wspierają hipotezę „Pożegnania z Afryką” (zob. Prehistoryczne wędrówki ludzkości), zgodnie z którą człowiek współczesny wyewoluował w Afryce i stamtąd kolejnymi falami migracji zaludniał Ziemię. Pokazują jednak historię zaledwie jednego fragmentu naszego genomu.
Genom
Mitochondria nazywane są autonomicznymi (bądź półautonomicznymi), gdyż są jednymi z niewielu organelli, które mają własny genom (syntezują około 10% białek). Ludzki genom mitochondrialny to kołowa cząsteczka DNA wielkości około 16 tysięcy par zasad. Koduje on 37 genów: 13 odpowiedzialnych za podjednostki kompleksów oddechowych I, III, IV i V, 22 kodujące mitochondrialne tRNA oraz dwa odpowiedzialne za rRNA. Jedno mitochondrium może zawierać od dwóch do dziesięciu kopii jego DNA.
Tak jak u prokariotów, w genomie mitochondrialnym występuje proporcjonalnie dużo kodującego DNA w stosunku do obszarów niekodujących oraz brak powtórzeń. Geny mitochondrialne są transkrybowane w postaci nici mRNA zawierającej kilka genów, która poddawana jest poliadenylacji w procesie obróbki posttranskrypcyjnej. Nie wszystkie białka niezbędne do działania mitochondriów kodowane są przez genom mitochondrialny. Większość z nich kodowana jest przez genom jądrowy, skąd transportowane są do mitochondriów. Dokładna liczba genów kodowanych przez genom jądrowy i mitochondrialny różni się w zależności od gatunku. Mitochondrialny DNA zwykle jest kołowy, chociaż zanotowano wyjątki od tej zasady. Ludzki mitochondrialny DNA pozbawiony jest także intronów, jednakże introny obecne są w genomie mitochondrialnym pewnych innych eukariontów, na przykład drożdży czy protistów takich jak Dictyostelium discoideum.
U zwierząt genom mitochondrialny jest zwykle pojedynczą, kołową cząsteczką DNA o wielkości 16 tysięcy par zasad, zawierającą 37 genów. Jednym z ciekawszych wyjątków jest genom mitochondrialny wszy ludzkiej. Jest on zorganizowany w postaci osiemnastu kołowych cząsteczek DNA, z których każda ma wielkość od trzech do czterech tysięcy par zasad i zawiera od jednego do trzech genów. Stwierdzono, że pomiędzy tymi cząsteczkami dochodzi do rekombinacji genetycznej.
O ile drobne różnice w odczytywaniu standardowego kodu genetycznego zostały przewidziane wcześniej, o tyle żadne nie zostały odkryte do 1979 roku, kiedy to uczeni badający ludzki genom mitochondrialny odkryli, że stosuje on inne od standardowego kodowanie. Od tamtej pory odkryto wiele takich drobnych różnic, w tym także różne rodzaje kodowania w genomach mitochondrialnych.
| Organizm | Kodon | Kodowanie standardowe | Kodowanie w mitochondriach |
|---|---|---|---|
| Ssaki | AGA, AGG | arginina | kodon stop |
| AUA | izoleucyna | metionina | |
| UGA | kodon stop | tryptofan | |
| Bezkręgowce | AGA, AGG | arginina | seryna |
| AUA | izoleucyna | metionina | |
| UGA | kodon stop | tryptofan | |
| Drożdże | AUA | izoleucyna | metionina |
| UGA | kodon stop | tryptofan | |
| CUA | leucyna | treonina |
Mutacje w genach mitochondrialnych powodują choroby mitochondrialne, których objawy dotykają głównie tkanki o największym zapotrzebowaniu energetycznym – mięśniową i nerwową. Choroby te mają charakterystyczny, matczyny wzór dziedziczenia. Również mutacje kodowanych w jądrze komórkowym białek mitochondrialnych powodują choroby genetyczne (np. ataksja Friedreicha).
Mitochondrialny DNA jest narażony na uszkodzenia przez wolne rodniki z łańcucha oddechowego, a w mitochondriach nie ma sprawnych mechanizmów naprawczych dla DNA. Leży to u podłoża hipotezy tłumaczącej objawy starzenia się akumulacją mutacji somatycznych mitochondrialnego DNA i obniżaniem sprawności energetycznej komórek.
Tak zwany obszar hiperzmienny mitochondrialnego DNA to niekodujący fragment genomu mitochondrialnego, który bardzo się różni między ludźmi. Dlatego wykorzystuje się go do badań genetyki populacyjnej oraz w medycynie sądowej do ustalania tożsamości. Sekwencje niektórych genów mitochondrialnych, różniące się między gatunkami, mogą służyć jako „kod kreskowy” charakterystyczny dla poszczególnych gatunków i są w związku z tym wykorzystywane w badaniach bioróżnorodności.
Ponieważ komórka zawiera tysiące kopii mitochondrialnego DNA, ma on większą szansę niż DNA jądrowy zachować się w materiale kopalnym. Do niedawna jedyne znane sekwencje kopalnego DNA były sekwencjami mitochondrialnymi. Porównanie sekwencji DNA mitochondrialnego ludzi współczesnych i neandertalczyków sugeruje, że gatunki te nie krzyżowały się.
Zobacz też
Bibliografia
- Eldra P. Solomon, Linda R. Berg, Diana W. Martin: Biologia. Warszawa: MULTICO Oficyna Wydawnicza, 2007. ISBN 978-83-7073-412-1. OCLC 177294444.
Linki zewnętrzne
- Mapa genomu ludzkiego mitochondrium na mitowheel.org