
Staw skroniowo-żuchwowy, staw żuchwowy wtórny (articulatio temporomandibularis) – występujący u ssaków staw łączący żuchwę z pozostałą częścią czaszki, dokładniej zaś z kością skroniową (lub łuskową). Należy do cech diagnostycznych ssaków.
Przodkowie ssaków posiadali pierwotny staw żuchwowy budowany przez kości kwadratową i stawową, a ich żuchwę budowały liczne kości. W trakcie ewolucji kości kwadratowa i stawowa stały się kosteczkami słuchowymi, rolę żuchwy pełnić zaczęła pojedyncza kość zębowa, która utworzyła wtórny staw żuchwowy, czyli staw skroniowo-żuchwowy, z kością skroniową bądź łuskową (uznawaną często za część kości skroniowej). Staw ten powstaje z komórek ektomezenchymalnego grzebienia nerwowego. Główkę stawową tworzy jeden z wyrostków kości zębowej, panewkę zaś kość skroniowa. Jamę stawową dzieli krążek stawowy. Zakres ruchów zachodzących w stawie zależy od taksonu, u niektórych możliwe są ruchy w trzech płaszczyznach umożliwiające żucie.
Pierwotny staw żuchwowy bazalnych czworonogów

jaw joint – staw żuchwowy
squamosal – kość łuskowa
dentary – kość zębowa
quadrate – kość kwadratowa
articular – kość stawowa
angular – kość kątowa
Staw skroniowo-żuchwowy nie występuje u płazów i gadów. Cechują się one inną budową anatomiczną. Posiadają one staw żuchwowy pierwotny. Ich kość zębowa stanowi tylko jedną z kilku kości żuchwy i nie bierze ona udziału w tworzeniu stawu żuchwowego. W skład stawu żuchwowego pierwotnego wchodzą bowiem kość kwadratowa (czworoboczna) i kość stawowa.
Pierwotne czworonogi odbierały drgania podłoża poprzez kończyny, przenoszące je dalej przez kości skórne łuku szczękowego do puszki mózgowej. Na lądzie okazało się to skutecznym sposobem odbierania informacji z otoczenia. Usprawniło to jeszcze wytworzenie się błony bębenkowej i strzemiączka, przekazujących drgania do ucha wewnętrznego. U płazów błona bębenkowa leży właśnie za stawem żuchwowym pierwotnym, niedaleko kości kwadratowej, uczestniczącej w przekazywaniu drgań, co umożliwiła ruchliwość szczęki.
Budowa pierwotnego stawu żuchwowego gadów pozwala jedynie na otwieranie i zamykanie jamy gębowej. Dlatego gady nie żują. Ruchy takie zapewniają mięsień skrzydłowy zginający żuchwę w sposób niosący za sobą ryzyko zwichnięcia oraz mięsień skroniowy. Duże siły działające na kość kwadratową wymagają też jej silnego połączenia z resztą czaszką, kostnego bądź dzięki tkankom miękkim, jak to ma miejsce u współczesnych lepidozaurów.
Zmiany u gadów ssakokształtnych
U gadów ssakokształtnych błona bębenkowa przeniosła się jeszcze niżej, a kość stawowa została włączona w łańcuch przekazujący drgania dźwiękowe. W efekcie kości tworzące aparat gębowy zaangażowały się w przewodzenie drgań. Wiązało się to z chrzęszczeniem w uszach podczas jedzenia.
Kość zębowa synapsydów w trakcie ewolucji powiększała się. Bardziej pierwotny pelykozaur Haptodus ma jeszcze żuchwę złożoną z wielu kości i staw żuchwowy pierwotny, podobnie jak terapsyd Biarmosuchus. Jednakże u takich cynodontów jak procynozuch widać już znacznie powiększoną kość zębową.
Zmiany dokonały się także w umięśnieniu. Doły skroniowe już wcześniej uległy otwarciu, (co widać u Petrolacosaurus czy Archaeothyris), co umożliwiło ekspansję przyczepów mięśni, mieszczących się w otworach podczas skurczu. Synapsydy cechowały się dużymi mięśniami poruszającemu żuchwą. Dopiero u tej grupy przyczepy mięśni poruszających żuchwę przeniosły się na zewnętrzną jej powierzchnię, jak to miało miejsce u terapsydów. Znalazł się tam przyczep żwacza, początkowo mięśnia niewielkich rozmiarów, przebiegającego z żuchwy do łuku jarzmowego, o działaniu antagonistycznym do mięśni skroniowego i skrzydłowego (do góry i zewnętrznie). Z biegiem czasu żwacz powiększał się, powiększała się i kość zębowa, do której się przyczepiał. Powiększona kość zębowa występuje u cynodontów. Natomiast w górnej części żuchwy powstał grzebień stanowiący miejsce przyczepu mięśnia skroniowego. Odtąd dwa mięśnie o przeciwnym działaniu razem zapewniały możliwość żucia dzięki ruchom w kierunku bocznym. Początkowo taki sposób rozdrabniania pokarmu utrudniał fakt uczestniczenia wielu kości w budowie żuchwy; łączyły je szwy, co narażało synapsyda na niebezpieczeństwo przerwania tych połączeń, zwłaszcza podczas wzrostu osobniczego, tym bardziej że podczas żucia działają duże siły.
Równolegle z powiększaniem się kości zębowej musiały zmniejszać się inne kości tworzące żuchwę. U cynodontów kość kwadratowa jest już niewielka i zagłębiona w kości łuskowej.
W dalszym przebiegu ewolucji postępujący rozrost kości zębowej sprawił, że sięgnęła ona aż do stawu żuchwowego, tworzonego dotychczas przez kości kwadratową i stawową. Kość zębowa również zaczęła uczestniczyć w tworzeniu połączenia stawowego z kością skroniową, najpierw obok starszego połączenia. Tak więc obok siebie funkcjonowały dwa parzyste połączenia żuchwy z resztą czaszki. Stan taki obserwuje się u zaawansowanych ewolucyjnie cynodontów z triasu środkowego i późnego. Jako przykład służyć może tutaj Probainognathus, u którego kość zębowa za pomocą wyrostka swej ogonowej części łączyła się z kością łuskową stawowo poprzez kości stawową i nadkątową. Natomiast żyjący na przełomie triasu i jury Morganucodon, również posiadający podwójny staw żuchwowy, nosił na tworzącej prawie samodzielnie żuchwę kości zębowej wyrostek kłykciowy, łączący się z kością łuskową. Rozwiązanie takie bardzo przypominało spotykane u ssaków. Zachował się również u niego, choć słabo wykształcony, pierwotny staw żuchwowy, jednak tworzące go kości: kwadratowa (zawieszona na kości łuskowej) i stawowa znajdują się już w okolicy ucha, przyśrodkowo do stawu wtórnego, tworząc przy współpracy ze strzemiączkiem łańcuch kosteczek słuchowych podobny do spotykanego obecnie u ssaków. Natomiast jego żuchwę budowała głównie kość zębowa, której towarzyszyło 6 innych, wysoce uwstecznionych kości. Cechy morganukodontów uważa się za najbardziej zaawansowane ewolucyjnie wśród terapsydów. Podczas gryzienia ich żuchwa poruszała się w kierunku grzbietowo-przyśrodkowym, jak u ssaków. Istniały też pewne rodziny cynodontów (Transversodontidae, Cynognathidae, Chiniquodontidae), u których wykształcił się wtórny kontakt pomiędzy kośćmi łuskową a surangular, blokujący nadmierne ruchy żuchwy w tył. U późniejszych ssaków rolę tę przejmie wyrostek zapanewkowy. Probainognathus z rodziny Chiniquodontidae wykształcił dół panewkowy prawdopodobnie z chrząstką stawową i mazią. Występowało połączenie stawowe z surangular. Obok niej znajdowała się podpierająca ją struktura przywodząca na myśl wyrostek stawowy kości zębowej, mogąca wstępować w obręb torebki stawowej. U niektórych morganukodontów kość zębowa rozrosła się jeszcze do tyłu, tracąc połączenie z surangular. Inne rozwiązanie rozwinęły Ictidosauria, u których staw tworzył kłykieć żuchwy, a dół panewkowy nie występował, chyba że panewkę tworzyła w całości nie zachowana w zapisie kopalnym tkanka chrzęstna. Równolegle z opisanymi zmianami u cynodontów rozwijały się mięśnie żwacze. Ruchy poprzeczne żuchwy poddane zostały dokładniejszej kontroli z jednej strony, a siła ugryzienia zwiększyła się z drugiej. Co więcej, redukcji uległy napięcia, na które narażone były stawy żuchwy.
Morganukodonty nie były jedynymi synapsydami posiadającymi obok siebie dwa parzyste stawy żuchwowe. Cechę taką zaobserwowano również u Kuehneotheriidae, takich jak niewielkie owadożerne Kuehneotherium o dobrze rozwiniętym kłykciu kości zębowej, zredukowanych kościach zazębowych. Natomiast budową zębów i innymi cechami różniło się ono od morganukodontów. Dwukrotne i jednoczesne w dodatku pojawienie się tej samej cechy (w tym wypadku podwójnego stawu żuchwowego) nie jest niemożliwe, choć w opisach ewolucji preferuje się zwykle najprostsze wyjaśnienie, a więc zakładające najmniej homoplazji. Dwukrotne pojawienie się stawu żuchwowego wtórnego stawia jednak pod znakiem zapytania monofiletyzm ssaków. Jeśli zaś omawiana cecha w rzeczywistości rozwinęła się u wspólnego przodka tych dwu grup (a morganukodonty i Kuehneotheriidae mogą być blisko spokrewnione), monofiletyzm będzie zachowany pod warunkiem włączenia rzeczonego przodka do grupy.
Tak więc w końcu staw pomiędzy kośćmi kwadratową i stawową przestał odgrywać istotną rolę w łączeniu żuchwy z resztą czaszki, zachowując jednak swą wagę w przenoszeniu drgań dźwiękowych. Ich połączenia z żuchwą zanikły, w przeciwieństwie do połączeń ze strzemiączkiem. W końcu przeniosły się one do ucha środkowego (przewodzenie dźwięków stało się ich główną funkcją i dzięki utracie powiązań z żuchwą mogły robić to skuteczniej, skuteczniej również można było dzięki temu przeżuwać pokarm). Powstał staw żuchwowy wtórny tworzony przez kość zębową i kość skroniową. Kość zębowa jest jedyną kością żuchwy. Stan taki obserwuje się u wczesnojurajskiego Hadrocodium, żyjącego przed 195 milionami lat na terenie współczesnych Chin. U rodzaju tego staw ten położony jest też trochę inaczej: do przodu od okienka owalnego i kłykci potylicznych. Za stawem leży dobrze rozwinięta okolica zapanewkowa z zagłębieniem na bocznej części kości łuskowej. Podobne cechy występowały u trikonodontów, wielu multituberkulatów, Obdurodon i Tachyglossus zaliczanych do stekowców, spotyka się je również u większości ssaków żyworodnych. U Hadrocodium dodatkowo puszka mózgowa i podstawa czaszki były względem stawu przesunięte w tył. Jest to także pierwszy znany takson, u którego kostki słuchowe utraciły już swe połączenie z żuchwą.
Jeszcze dzisiaj staw ten wykształca się u ssaków po okresie ssania.
W przeszłości uważano, że taka przemiana nie jest możliwa. Podawano ją nawet jako argument przeciwko ewolucji. Obecnie procesy te, związane z powstaniem ssaków, uważa się za jedne z najlepiej udokumentowanych przykładów powstawania dużych taksonów. Zmiany prowadzące do powstania ssaków z gadów zachodziły stopniowo, bez nagłych skoków.
Embriologia
Komórki uczestniczące w morfonegezie czaszki pochodzą z okolicy głowowej części grzebienia nerwowego. Komórki z okolic drugiego i trzeciego pęcherzyka pierwotnego migrują w kierunku tworzących się łuków skrzelowych. Staw żuchwowy rozwija się z pierwszego z łuków skrzelowych.
By komórki ektomezenchymalne grzebienia nerwowego po migracji mogły stać się kością czy chrząstką, muszą przejść zmiany indukcyjne. Odgrywają w nich rolę liczne czynniki transkrypcyjne. U myszy czy danio w powstaniu kości żuchwy istotną rolę odgrywa endotelina 1. Myszy pozbawione genu receptora endoteliny A wykazywały defekty żuchwy, w tym nieprawidłowe połączenia z innymi kośćmi, defekty wykazały też zwierzęta pozbawione genu sucker kodującego samą endotelinę bądź też genu Ece-1 kodującego enzym konwertujący endotelinę. Endotelinę 1 wydzielają komórki rdzennej mezodermy przyosiowej i otaczającego ją nabłonka ekto- i endodermalnego. Oddziałuje ona na receptory komórek pochodzących z grzebienia nerwowego, którego odbyły już migrację. Dzięki niej następuje zróżnicowanie w osi grzbietowo-brzusznej. Mutanty pozbawione suckwer wykazują też nieprawidłowości w ekspresji dHAND, dlx2, msxE, gsc, dlx3 i EphA3. Najważniejszym efektorem wydaje się hand2.
Ważną rolę w tworzeniu się stawu żuchwy odgrywa również gen Bapx1 z rodziny czynników transkrypcyjnych NK-2, homolog należącego do genów hemeobox muszki owocowej bagpipe. U ryb (danio pręgowany) brak stawu żuchwowego może też mieć związek z nadeksrepsją bapx1. Wśród ssaków sklonowano go u myszy. Występuje też u innych kręgowców, u których – regulowany przez endotelinę 1 – bierze udział w tworzeniu pierwotnego stawu żuchwy, ulegając ekspresji w mezenchymie twarzowej. Gen ten u płazów prócz jelita ulega ekspresji w chrząstce budującej twarz. Wydaje się on mieć związek z tworzeniem się osi prawo-lewo, gdyż jego ekspresja u ssaków i ptaków (ptasi homolog nazywa się NKX3.2) wykazuje asymetrię, co więcej, o ile u ptaków (kurczak) ekspresja przeważała w mezodermie bocznej po stronie lewej, to u ssaków (mysz) przeważała strona prawa – już od dziewiątego dnia licząc od stosunku, początkowo w somitach, potem w mezodermie bocznej. Gen ten wpływa poza tworzeniem się stawu żuchwowego również na rozwój typowego dla ssaków ucha środkowego z trzema kosteczkami słuchowymi.
Nieprawidłowości żuchwy wykazywały też mysie noworodki goosecoid-null. Natomiast nie biorą w tym procesie udziału geny Hox, uczestniczące w tworzeniu łuków skrzelowych od 2 do 6. W tworzeniu chrząstki odgrywa też rolę kontakt komórek grzebieni nerwowych z endodermą, ektoderma zaś hamuje taką przemianę.
Anatomia
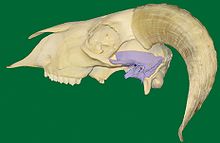

Natomiast u ssaków żuchwę buduje pojedyncza parzysta kość zębowa, zrastająca się z przeciwstronną kością jednoimienną chrząstkozrostem międzyżuchwowym, tworząca staw z łuską kości skroniowej za pomocą swego wyrostka kłykciowego przechodzącego w głowę żuchwy. Z kolei kość skroniowa wytwarza takie struktury, jak guzek stawowy, dół żuchwowy i wyrostek zastawkowy. Dalsze szczegóły budowy stawu zależą już bardzo od gatunku, zwłaszcza zaś od rodzaju spożywanego przez dany gatunek pożywienia.
Pierwotnie uznawano opisywany staw za niekontaktujący, bowiem powierzchnie stawowe kości skroniowej i żuchwy nie były do siebie dobrze dopasowane. Obecnie pogląd ten jest inny: staw określa się jako kontaktujący ze względu na istnienie pomiędzy powierzchniami stawowymi krążka stawowego z chrząstki włóknistej, pogrubionego na obwodzie i zrastającego się z torebką stawową, dzielącego jamę stawową na piętro górne i dolne.
Torebka stawu żuchwowego wtórnego wzmacniana jest przez więzadło boczne oraz więzadło doogonowe. To ostatnie przebiega od wyrostka zastawowego kości skroniowej do wyrostka kłykciowego kości zębowej.
Krążek stawowy u kopytnych jest grubszy, niż u drapieżnych.
Koń posiada żółte, elastyczne więzadło doogonowe.

U człowieka powierzchnia stawowa budowana jest przed przednią część dołu żuchwowego (tylną wyściela tkanka łączna, w tym tkanka tłuszczowa), dokładniej zaś dołek stawowy, leżący dziobowo od szczeliny skalisto-bębenkowej, oraz guzek stawowy w kształcie poprzecznego wału. Powierzchnię stawową pokrywa chrząstka włóknista. Z kolei głowa żuchwy przybiera kształt elipsoidalny, z powierzchnią stawową skierowaną do góry i dziobowo. Krążek stawowy człowieka to grubsza na obwodzie płytka mierząca 3-4 mm, odtwarzająca kształt powierzchni stawowych. Luźna torebka stawowa ma kształt lejka. Obejmuje z jednej strony głowę żuchwy aż do szyjki u dołu (przyczyną większego zasięgu torebki z tyłu niż z przodu jest pozostający z konieczności na zewnątrz jamy stawowej przyczep mięśnia skrzydłowego bocznego), z drugiej guzek i dołek stawowy do szczeliny skalisto-bębenkowej. Występują dwie odrębne błony maziowe w górnym i dolnym piętrze stawu. Ponadto wyróżnia się więzadło boczne (skroniowo-żuchwowe) o skośnym przebiegu, dobrze zbudowane i trójkątne w kształcie, więzadło klinowo-żuchwowe leżące przyśrodkowo oraz więzadło rylcowo-żuchwowe sięgające wyrostka rylcowatego.
Ogólnie staw ten jest plastyczny, dopasowuje się do uzębienia, do zgryzu.
U ludzkiego noworodka nie ma jeszcze guzka stawowego. Płaski dół żuchwowy uwypukla się dopiero wraz z rozwojem zębów. Natomiast u starszych ludzi, pozbawionych uzębienia, spłaszczają się i guzek, i dołek stawowy. Krążek stawowy może w ogóle zaniknąć, co skutkuje opadaniem głowy żuchwy do panewki i ograniczeniem jej ruchu do ruchu zawiasowego.
Budowa opisywanego stanowi zależy także od rodzaju zwarcia. Zwarcie obcęgowe wiąże się z płaskim, saneczkowym stawem o kiepsko rozwiniętym guzku stawowym. Natomiast w zwarciu nożycowatym guzek jest dobrze rozwinięty, pochylony, natomiast przeważającym ruchem jest ruch zawiasowym. Staw ten dostosowuje się również do ubytków w uzębieniu.
Unaczynienie u człowieka zapewniają gałązki tętnicy szyjnej zewnętrznej, zwłaszcza poprzez tętnicę skroniową powierzchowną. Limfa odpływa do głębokich węzłów chłonnych szyjnych górnych. Unerwienie pochodzi od nerwów żwaczowego i uszno-skroniowego.
Działanie
Staw żuchwowy wtórny wraz z uzębieniem, mięśniami żwaczowymi i żuchwą tworzy jeden układ czynnościowy.
Staw ten klasyfikuje się jako staw kłykciowy. Działa jak zwykły staw zawiasowy, gdy żwacz unosi żuchwę, wykonuje zaś ruchy ślizgowe, gdy ciągnie go mięsień skroniowy. U człowieka ruchy w stawie przebiegają w trzech płaszczyznach: przód-tył, góra-dół oraz z boku na bok (ruchy boczne, żuciowe).
Ruchy zapewniają mięśnie żuciowe. Mięsień żwacz unosi kość zębową. Ruchy ślizgowe zapewnia mięsień skroniowy.
Staw o opisanej budowie umożliwia ruchy żuchwy w górę i w dół, pozwalające na zwarcie łuków zębowych. Co więcej, możliwe są również ślizgowe (saneczkowe) ruchy w kierunku dziobowo-ogonowym, które zapewnia obecność wspomnianego krążka w jamie stawowej. Symetryczne po obu stronach ruchy w tym kierunku pozwalają ssakom rozcierać i przegryzać. Psy wykorzystują je choćby do czyszczenia swego futra z pcheł. Istnieje także możliwość, by po obu stronach czaszki ruchy w rzeczonych stawach zachodziły w odwrotną stronę. W efekcie powstają ruchy ósemkowe żuchwy, dzięki którym przeżuwacze przeżuwają pokarm, bądź też żuchwa skręca bocznie, umożliwiając łamaczom drapieżnych kruszyć kości. Ruchy takie nie występują u części łasicowatych, którym uniemożliwia to szczelne objęcie wałeczkowatych głów kości zębowej przez kość skroniową. U drapieżnych ogólnie jest to staw zawiasowy dzięki ścisłemu objęciu poprzecznie położonego bloczka panewką. Działanie poruszających stawem mięśni u psów badał Dessem. Siła rozwijana podczas gryzienia przewyższała osiąganą podczas żucia. Podczas żucia mierzona elektromiograficznie aktywność mięśni żwaczy była niejednakowa – większa po stronie mięśnia odgrywającego w danej chwili główną rolę, mniejszą w mięśniu kontralateralnym, który nie osiągał takiej aktywności nawet podczas miażdżenia kości. Znaczna aktywność mięśnia przeciwstronnego nie byłaby bowiem korzystna z punktu widzenia mechaniki. Ważną rolę odgrywa też mięsień skroniowy. U świni również ważną rolę odgrywają te 2 mięśnie, które, przywodząc żuchwę dzięki przyczepom na zewnętrznych powierzchniach żuchwy i czaszki, zamykają usta. Natomiast w przypadku ruchów poprzecznych wymienione mięśnie jednoimienne działają antagonistycznie. Mięsień skroniowy ipsilateralnie pociąga żuchwę w tył, mięsień żwacz pociąga ją przeciwstronnie do przodu, tak więc prawy żwacz współpracuje z lewym mięśniem skroniowym i odwrotnie. Świnia wykonuje ruchy poprzeczne podczas żucia, jednostronnego u tego wszystkożercy. Wpierw następuje ruch w stronę, po której następuje aktywne żucie, następnie zaś w przeciwną.
U gryzoni dominują ruchy saneczkowe dzięki strzałkowemu ustawieniu walcowatej głowy żuchwy.
U człowieka krążek stawowy działa jako ruchoma panewka. Podczas otwierania jamy ustnej jest ona przemieszczana z dołka stawowego na guzek stawowy razem z głową żuchwy dzięki pracy mięśnia skrzydłowego bocznego. Analogiczny ruch głowy żuchwy umożliwia luźność torebki stawowej. W spoczynku szeregi zębów nie ulegają całkowitemu zamknięciu na skutek nieścisłego przylegania. Głowa żuchwy znajduje się na powierzchni tylnej guzka stawowego, rozdzielona odeń częścią krążka. Sam dołek stawowy wypełnia tylna część krążka. Głowa może dostawać się do dołka podczas przechylenia głowy w tył, na przykład w czasie snu. U człowieka ruchy w obu stawach (po stronie prawej i lewej) zawsze odbywają się równocześnie, wobec czego mówi się o jednym łańcuchu obejmującym 4 jamy stawowe. Jednakże u człowieka ruchy w omawianych stawach są wielostronne, podczas gdy u większości ssaków są one prostsze. Tak więc człowiek dokonuje w tym stawie:
- ruchów saneczkowych, czyli ślizgowych polegających na wysuwaniu lub cofaniu kości zębowej. Podczas takiego ruchu głowa żuchwy ześlizguje się razem z krążkiem z dołka stawowego, posuwając się po powierzchni tylnej guzka stawowego w kierunku przednio-dolnym, dążąc ku szczytowi wspomnianego guzka, który z kolei oddala się od guzka przeciwstronnego. U ludzi chudych opisany ruch głowy żuchwy można zaobserwować – między głową żuchwy a małżowiną uszną tworzy się zagłębienie. Podczas takiego ruchu siekacze dolne zajmują miejsce przed górnymi. Odwrotne ruchy mają miejsce, gdy żuchwa wraca.
- ruchów zawiasowych umożliwiających mu otwieranie i zamykanie ust poprzez podnoszenie czy też opuszczanie żuchwy. W górnym piętrze stawu również zachodzi ruch ślizgowy, podczas gdy w dolnym zachodzi ruch zawiasowy, jednoczesny z tamtym. Głowa żuchwy z dołka, w którym przebywa w spoczynku, przemieszcza się ruchem ślizgowym na guzek, dokonując obrotu na dolnej powierzchni krążka stawowego wokół osi poprzecznej przecinającej głowę żuchwy. W efekcie zęby dolne oddalają się od górnych, usta otwierają się. Odwrotnie przebiega ruch zamykania ust.
- w końcu zaś ruchów żucia, czyli mielenia, będących przemieszczeniami kości zębowej na boki. Następuje obrót kości zębowej wokół osi ustawionej pionowo przechodzącej przez jedną z głów żuchwy (zachowują się one podczas tego ruchu niejednakowo). Bródka zbliża się też do jednej z głów kości zębowej, tej pozostającej w dołku. Szeregi zębów oddalają się wtedy od siebie na wysokość guzka, może znaleźć się między nimi kęs pokarmu. Dalej następuje zmiana strony (jedna z głów, się cofa, a druga wysuwa).
Człekokształtne wyróżniają się tutaj brakiem specjalizacji w kierunku jednego z tych ruchów.
Patologia
Opisano różne schorzenia związane ze stawem skroniowo-żuchwowym. U człowieka wymieniane są:
- ankyloza, leczona operacyjnie, mogąca wynikać ze zmian ropnych, pooperacyjnych, nowotworowych, zapalnych, po radioterapii
- chondromatoza synowialna
- przemieszczenie krążka, wiążące się z remodelingiem tkanek miękkich stawu
- przemieszczenie żuchwy
- stany zapalne, w tym artropatia łuszczycowa, reumatoidalne zapalenie stawów, zesztywniające zapalenie stawów kręgosłupa
- nowotwory, w tym osteochondroma
- urazy
- zmiany degeneracyjne
Bibliografia
- Michał Reicher, Wiesław Łasiński: Połączenia stawowe. Staw skroniowo-żuchwowy. W: Adam Bochenek, Michał Reicher, Tadeusz Bilikiewicz, Stanisław Hiller, Eugenia Stołyhwo: Anatomia człowieka. T. I: Anatomia ogólna. Kości. Stawy i więzadła. Mięśnie. Warszawa: Wydawnictwo Lekarskie PZWL, 2007. ISBN 978-83-200-3682-4.
- Jerzy Dzik: Dzieje życia na Ziemi. Warszawa: Wydawnictwo Naukowe PWN, 2011. ISBN 978-83-01-16678-6.
- Douglas J. Futuyma: Ewolucja. Warszawa: Wydawnictwa Uniwersytetu Warszawskiego, 2008. ISBN 978-83-235-0577-8.
- Kazimierz Krysiak, Henryk Kobryń, Franciszek Kobryńczuk: Anatomia zwierząt. T. 1: Aparat ruchowy. Warszawa: Wydawnictwo Naukowe PWN, 2012. ISBN 978-83-01-16755-4.
- A. W. Crompton, Farish A. Jenkins: Origin of Mammals. W: Jason A. Lillegraven, Zofia Kielan-Jaworowska, William A. Clemens: Mesozoic Mammals: The First Two-Thirds of Mammalian History. University of California Press, 1979.